Зоология беcпозвоночных
Класс Головоногие (Cephalopoda)
Головоногие - самые высокоорганизованные моллюски. Их справедливо называют "приматами" моря среди беспозвоночных животных за совершенство их приспособлений к жизни в морской среде и сложность поведения. Это в основном крупные хищные морские животные, способные активно плавать в толще воды. К ним относятся кальмары, осьминоги, каракатицы, наутилусы (рис. 234). Их тело состоит из туловища и головы, а нога преобразована в щупальца, расположенные на голове вокруг рта, и особую двигательную воронку на брюшной стороне тела (рис. 234, А). Отсюда произошло название - головоногие. Доказано, что часть щупалец головоногих образуется за счет головных придатков.
У большинства современных головоногих раковина отсутствует или рудиментарна. Только у рода наутилус (Nautilus) имеется спирально закрученная раковина, разделенная на камеры (рис. 235).
К современным головоногим относится всего 650 видов, а ископаемых видов насчитывают около 11 тыс. Это древняя группа моллюсков, известная с кембрия. Вымершие виды головоногих были преимущественно раковинными и имели наружную или внутреннюю раковину (рис. 236).
Для головоногих характерны многие прогрессивные черты организации в связи с активным образом жизни морских хищников. Вместе с тем у них сохраняются некоторые примитивные признаки, свидетельствующие об их древнем происхождении.
Внешнее строение. Особенности внешнего строения головоногих разнообразны в связи с различным образом жизни. Их размеры колеблются от нескольких сантиметров до 18 м у некоторых кальмаров. Нектонные головоногие обычно торпедовидной формы (большинство кальмаров), бентосные имеют мешковидную форму тела (многие осьминоги), нектобентосные - уплощенную (каракатицы). Планктонные виды мелкие по размерам, имеют студенистое плавучее тело. Форма тела у планктонных головоногих может быть узкой или похожей на медуз, а иногда шаровидной (кальмары, осьминоги). Бентопелагические головоногие обладают раковиной, поделенной на камеры.
Тело головоногих состоит из головы и туловища. Нога модифицирована в щупальца и воронку. На голове расположен рот, окруженный щупальцами, и крупные глаза. Щупальца образованы головными придатками и ногой. Это органы захвата пищи. У примитивного головоногого - кораблика (Nautilus) щупалец неопределенное количество (около 90); они гладкие, червеобразные. У высших головоногих щупальца длинные, с мощной мускулатурой и несут крупные присоски на внутренней поверхности. Число щупалец 8- 10. У головоногих с 10 щупальцами два щупальца - ловчие, более длинные, с присосками на расширенных концах,
322

Рис. 234. Головоногие моллюски: А - наутилус Nautilus, Б - осьминог Benthoctopus; 1 - щупальца, 2 - воронка, 3 - капюшон, 4 - глаз
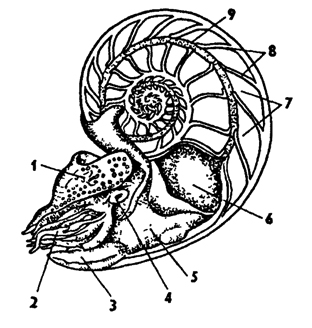
Рис. 235. Наутилус Nautilus pompilius с распиленной раковиной (по Оуэну): 1 - головной капюшон, 2 - щупальца, 3 - воронка, 4 - глаз, 5 - мантия, 6 - внутренностный мешок, 7 - камеры, 8 - перегородка между камерами раковины, 9 - сифон
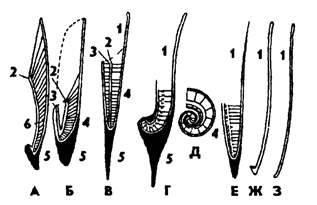
Рис. 236. Схема строения раковин головоногих в саггитальном разрезе (из Гешелера): А - Sepia, Б - Belosepia, В - Belemnites, Г - Spirulirostra, Д - Spirula, Е - Ostracoteuthis, Ж - Ommastrephes, З - Loligopsis (В, Г, Е - ископаемые); 1 - проостракум, 2 - спинной край сифональной трубки, 3 - брюшной край сифональной трубки, 4 - совокупность камер-фрагмокон, 5 - рострум, 6 - полость сифона
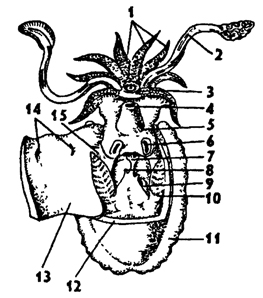
Рис. 237. Мантийная полость каракатицы - Sepia (по Пфуршеллеру): 1 - короткие щупальца, 2 - ловчие щупальца, 3 - рот, 4 - отверстие воронки, 5 - воронка, 6 - хрящевые ямки запонок, 7 - анус, 8 - почечные сосочки, 9 - половой сосочек, 10 - жабры, 11 - плавник, 72 - линия отреза мантии, 13 - мантия, 14 - хрящевые бугорки запонок, 15 - мантийный ганглий
323
а остальные восемь щупалец более короткие (кальмар, каракатица). У осьминогов, обитающих на морском дне, восемь щупалец одинаковой длины. Они служат осьминогу не только для захвата пищи, но и для передвижения по дну. У самцов осьминогов одно щупальце видоизменено в половое (гектокотиль) и служит для переноса половых продуктов в мантийную полость самки.
Воронка - производное ноги у головоногих, служит для "реактивного" способа движения. Через воронку вода с силой выталкивается из мантийной полости моллюска, и его тело движется реактивно в противоположном направлении. У кораблика воронка не срослась на брюшной стороне и напоминает свернутую в трубку подошву ноги ползающих моллюсков. Доказательством того, что щупальца и воронка головоногих - производные ноги, служит их иннервация от педальных ганглиев и эмбриональная закладка этих органов на брюшной стороне зародыша. Но, как уже отмечалось, часть щупалец головоногих - производные головных придатков.
Мантия на брюшной стороне образует как бы карман - мантийную полость, открывающуюся наружу поперечной щелью (рис. 237). Из этой щели выступает воронка. На внутренней поверхности мантии имеются хрящевые выступы - запонки, которые плотно входят в хрящевые углубления на теле моллюска, и мантия как бы пристегивается к телу.
Мантийная полость и воронка в совокупности обеспечивают реактивное движение. При расслаблении мускулатуры мантии вода входит через щель в мантийную полость, а при ее сокращении полость закрывается на запонки и вода выталкивается через воронку наружу. Воронка способна изгибаться вправо, влево и даже назад, что обеспечивает разное направление движения. Роль руля дополнительно выполняют щупальца и плавники - кожные складки туловища. Типы движения у головоногих разнообразны. Осьминоги чаще передвигаются на щупальцах и реже плавают. У каракатиц кроме воронки для движения служит круговой плавник. Некоторые глубоководные осьминоги зонтикообразной формы имеют перепонку между щупальцами - умбреллу и могут передвигаться за счет ее сокращений, подобно медузам.
Раковина у современных головоногих рудиментарна или отсутствует. У древних вымерших головоногих раковина была хорошо развита. Только один современный род Nautilus сохранил развитую раковину. Раковина Nautilus и у ископаемых форм обладает существенными морфофункциональными особенностями, в отличие от раковин других моллюсков. Это не только защитное приспособление, но и гидростатический аппарат. У наутилуса спирально закрученная раковина разделена перегородками на камеры. Тело моллюска размещается только в последней камере, открывающейся устьем наружу. Остальные камеры заполняются газом и камерной жидкостью, что обеспечивает плавучесть тела моллюска. Через
324
отверстия в перегородках между камерами раковины проходит сифон - задний отросток тела. Клетки сифона способны выделять газы. При всплывании моллюск выделяет газы, вытесняя камерную жидкость из камер; при опускании на дно моллюск заполняет камеры раковины камерной жидкостью. Движителем у наутилуса является воронка, а раковина поддерживает его тело во взвешенном состоянии в воде. Ископаемые наутилиды имели подобную раковину, как у современного наутилуса. У полностью вымерших головоногих - аммонитов также была наружная, спирально закрученная раковина с камерами, но у них перегородки между камерами имели волнистое строение, что увеличивало прочность раковины. Именно поэтому аммониты могли достигать очень крупных размеров, до 2 м в диаметре. У другой группы вымерших головоногих - белемнитов (Belemnoidea) раковина была внутренняя, обросшая кожей. Белемниты по внешнему облику напоминали безраковинных кальмаров, но в их туловище находилась коническая раковина, поделенная на камеры. Вершина раковины заканчивалась острием - рострумом. Рострумы раковин белемнитов часто встречаются в меловых отложениях, и их называют "чертовыми пальцами". У некоторых современных безраковинных головоногих имеются рудименты внутренней раковины. Так, у каракатицы на спине под кожей сохраняется известковая пластинка, имеющая на срезе камерное строение (238, Б). Только у спирулы (Spirula) под кожей находится вполне развитая спирально закрученная раковина (рис 238, А), а у кальмара под кожей сохранилась от раковины лишь роговая пластинка. У самок современных головоногих - аргонавтов (Argonauta) развита выводковая камера, напоминающая по форме спиральную раковину. Но это лишь внешнее сходство. Выводковая камера выделяется эпителием щупалец, очень тонкая и предназначена для защиты развивающихся яиц.
Покровы. Кожа представлена однослойным эпителием и слоем соединительной ткани. В коже имеются пигментные клетки - хроматофоры. Головоногим свойственна способность быстро изменять окраску. Этот механизм контролируется нервной системой и осуществляется за счет изменения формы
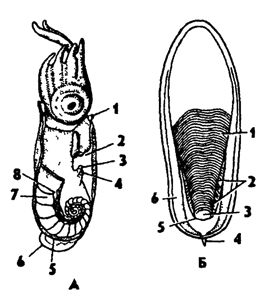
Рис. 238. Рудименты раковины у головоногих (по Натали и Догелю): А - спирула (Spirula); 1 - воронка, 2 - мантийная полость, 3 - анус, 4 - выделительное отверстие, 5 - орган свечения, 6 - плавник, 7 - раковина, 8 - сифон; Б - раковина Sepia; 1 - перегородки, 2 - боковой край, 3 - сифональная ямка, 4 - рострум, 5 - рудимент сифона, 6 - задний край проостракума
325
пигментных клеток. Так, например, каракатица, проплывая над песчаным грунтом, принимает светлую окраску, а над каменистым грунтом - темную. .При этом в ее коже пигментные клетки с темным и светлым пигментом попеременно то сжимаются, то расширяются. Если перерезать зрительные нервы у моллюска, то он теряет способность изменять окраску. За счет соединительной ткани кожи образуются хрящи: в запонках, основаниях щупалец, вокруг мозга.
Защитные приспособления. Головоногие, утратив в процесс эволюции раковину, приобрели другие защитные приспособления. Во-первых, от хищников многих из них спасает быстрое движение. Кроме того, они могут защищаться щупальцами и "клювом", представляющим собой видоизмененные челюсти. Крупные кальмары и осьминоги могут вступать в борьбу с крупными морскими животными, например с кашалотами. У малоподвижных и мелких форм развита покровительственная окраска и способность быстро изменять окраску. И наконец, у некоторых головоногих как, например, у каракатицы, имеется чернильный мешок, проток которого открывается в заднюю кишку. Выпрыскивание чернильной жидкости в воду вызывает как бы дымовую завесу, позволяющую моллюску скрыться от хищников в безопасное место. Пигмент чернильной железы каракатиц используется для изготовления высококачественной художественной туши.
Внутреннее строение головоногих
Пищеварительная система головоногих несет черты специализации к питанию животной пищей (рис. 239). Пищей им служат главным образом рыбы, крабы и двустворчатые моллюски. Добычу они схватывают щупальцами и убивают челюстями и ядом. Несмотря на крупные размеры, головоногие могут питаться только жидкой пищей, так как у них очень узкий пищевод, который проходит через мозг, заключенный в хрящевую капсулу. У головоногих имеются приспособления для перетирания пищи. Для разгрызания добычи им служат твердые роговые челюсти, похожие на клюв попугая. В глотке пища перетирается радулой и обильно смачивается слюной. В глотку впадают протоки 1-2 пар слюнных желез, которые выделяют ферменты, расщепляющие белки и полисахариды. Вторая задняя пара слюнных желез выделяет яд. Жидкая пища из глотки по узкому пищеводу поступает в энтодермальный желудок, куда впадают протоки парной печени, вырабатывающей разнообразные пищеварительные ферменты. Печеночные протоки усажены мелкими дополнительными железками, совокупность которых называют поджелудочной железой. Ферменты этой железы действуют на полисахариды,
326
и, следовательно, эта железа функционально отличается от поджелудочной железы млекопитающих. Желудок головоногих обычно со слепым мешковидным отростком, увеличивающим его объем, что позволяет им поглощать большую порцию пищи. Как и другие хищные животные, они едят много и относительно редко. От желудка отходит тонкая средняя кишка, которая потом переходит в заднюю, открывающуюся анальным отверстием в мантийную полость. В заднюю кишку у многих головоногих впадает проток чернильной железы, секрет которой имеет защитное значение.
Нервная система головоногих наиболее высокоразвитая среди моллюсков. Нервные ганглии образуют крупное окологлоточное скопление - мозг (рис. 240), заключенный в хрящевую капсулу. Имеются дополнительные ганглии. В состав мозга прежде всего входят: пара крупных церебральных ганглиев, иннервирующих голову, и пара висцеральных ганглиев, посылающих нервные тяжи к внутренним органам. По бокам от церебральных ганглиев расположены дополнительные крупные оптические ганглии, иннервирующие глаза. От висцеральных ганглиев отходят длинные нервы к двум мантийным ганглиям звездчатой формы, развивающиеся у головоногих в связи с функцией мантии в их реактивном способе движения. В состав мозга головоногих входят кроме церебральных и висцеральных педальные ганглии, которые подразделены на парные ганглии щупалец (брахиальные) и воронки (инфудибулярные). Примитивная нервная система, сходная с лестничной системой боконервных и моноплакофор, сохранилась только у Nautilus. Она представлена нервными тяжами, образующими окологлоточное кольцо без ганглиев и педальную дугу. Нервные тяжи покрыты нервными клетками. Такое строение нервной системы свидетельствует о древнем происхождении головоногих от примитивных раковинных моллюсков.
Органы чувств головоногих хорошо развиты. Особенно сложного развития у них достигают глаза, имеющие наибольшее значение для ориентации в пространстве и охоты за добычей. У Nautilus глаза имеют простое строение в виде глубокой глазной ямки (рис. 241, А), а у остальных головоногих глаза сложные - в форме глазного пузыря и напоминают строение глаза у млекопитающих. Это интересный пример конвергенции между беспозвоночными и позвоночными животными. На рисунке 241, Б изображен глаз каракатицы. Сверху глазное яблоко покрыто роговицей, в которой имеется отверстие в переднюю камеру глаза. Связь передней полости глаза с внешней средой предохраняет глаза головоногих от действия высокого давления на больших глубинах. Радужная оболочка образует отверстие - зрачок. Свет через зрачок попадает на шаровидный хрусталик, образованный эпителиальным телом - верхней оболочкой глазного пузыря. Аккомодация глаза у головоногих происходит иначе,
327
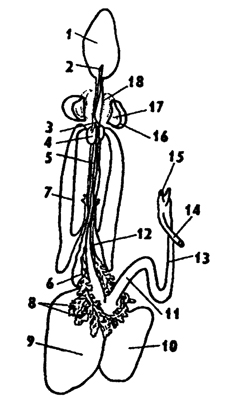
Рис. 239. Пищеварительная система каракатицы Sepia officinalis (по Резелеру и Лампрехту): 1 - глотка, 2 - общий слюнной проток, 3 - слюнные протоки, 4 - задняя слюнная железа, 5 - пищевод, 6 - головная аорта, 7 - печень, 8 - поджелудочная железа, 9 - желудок, 10 - слепой мешок желудка, 11 - тонкая кишка, 12 - печеночный проток, 13 - прямая кишка, 14 - проток чернильного мешка, 15 - анус, 16 - головная хрящевая капсула (разрезана), 17 - статоцист, 18 - нервное кольцо (разрезано)
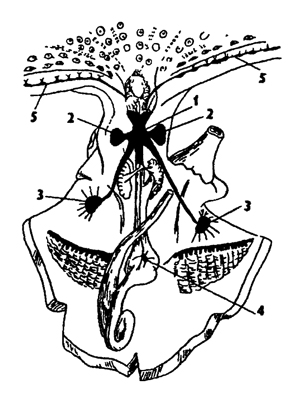
Рис. 240. Нервная система головоногих: 1 - мозг, 2 - оптические ганглии, 3 - мантийные ганглии, 4 - кишечный ганглий, 5 - нервные тяжи в щупальцах
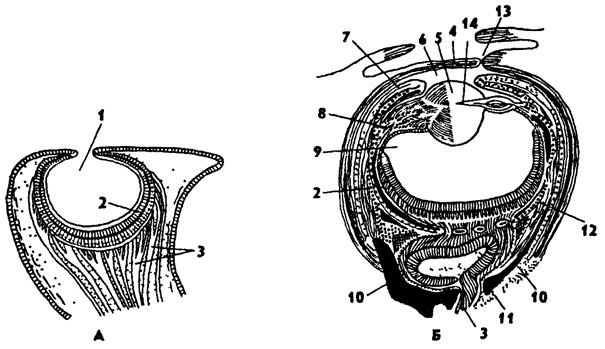
Рис. 241. Глаза головоногих: А - Nautilus, Б - Sepia (по Генсену); 1 - полость глазной ямки, 2 - сетчатка, 3 - зрительные нервы, 4 - роговица, 5 - хрусталик, 6 - передняя камера глаза, 7 - радужина, 8 - ресничный мускул, 9 - стекловидное тело, 10 - глазные отростки хрящевой капсулы, 11 - оптический ганглий, 12 - склера, 13 - отверстия камеры глаза, 14 - эпителиальное тело
328
чем у млекопитающих: не за счет изменения кривизны хрусталика, а путем его приближения или удаления от сетчатки (подобно фокусированию фотоаппарата). К хрусталику подходят особые ресничные мышцы, приводящие его в движение. Полость глазного яблока заполнена стекловидным телом, имеющим светопреломляющую фукнцию. Дно глаза выстлано зрительными - ретинальными и пигментными - клетками. Это сетчатка глаза. От нее отходит короткий зрительный нерв к оптическому ганглию. Глаза вместе с оптическими ганглиями окружены хрящевой капсулой. У глубоководных головоногих на теле имеются органы свечения, построенные по типу глаз.
Органы равновесия - статоцисты расположены в хрящевой капсуле мозга. Органы обоняния представлены обонятельными ямками под глазами или типичными для моллюсков осфрадиями у основания жабер - у наутилуса. Органы вкуса сосредоточены на внутренней стороне концов щупалец. Осьминоги, например, при помощи щупалец различают съедобные объекты от несъедобных. На коже головоногих множество осязательных и светочувствительных клеток. В поисках добычи они руководствуются сочетанием зрительных, осязательных и вкусовых ощущений.
Органы дыхания представлены ктенидиями. У большинства современных головоногих их два, а у наутилуса - четыре. Они расположены в мантийной полости по бокам туловища. Ток воды в мантийной полости, обеспечивающий газообмен, определяется ритмичным сокращением мускулатуры мантии и функцией воронки, через которую вода выталкивается наружу. Во время реактивного способа движения ток воды в мантийной полости ускоряется, а интенсивность дыхания возрастает.
Кровеносная система головоногих почти замкнутая (рис. 242). В связи с активным движением у них хорошо развиты целом и кровеносные сосуды и, соответственно, слабо выражена паренхиматозность. В отличие от остальных моллюсков, они не страдают гипокенией - слабой подвижностью. Скорость движения крови у них обеспечивается работой хорошо развитого сердца, состоящего из желудочка и двух (или четырех - у Nautilus) предсердий, а также пульсирующими участками сосудов. Сердце окружено обширной перикардиальной полостью,
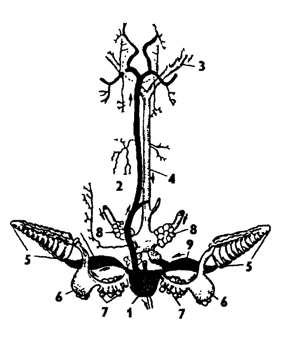
Рис. 242. Кровеносная система головоногих моллюсков (из Абрикосова): 1 - сердце, 2 - аорта, 3, 4 - вены, 5 - жаберные сосуды, 6 - жаберные сердца, 7, 8 - воротная система почек, 9 - жаберные вены
329
которая выполняет многие функции целома. От желудочка сердца отходят головная аорта - вперед и внутренностная аорта - назад. Головная аорта разветвляется на артерии, снабжающие кровью голову и щупальца. От внутренностной аорты отходят сосуды к внутренним органам. Кровь от головы и внутренних органов собирается в полую вену, расположенную продольно в нижней части туловища. Полая вена подразделяется на два (или четыре у Nautilus) приносящих жаберных сосуда, которые образуют сокращающиеся расширения - жаберные "сердца", способствующие жаберному кровообращению. Приносящие жаберные сосуды прилегают вплотную к почкам, образуя мелкие слепые впячивания в ткань почек, что способствует освобождению венозной крови от продуктов обмена. В жаберных капиллярах происходит окисление крови, которая затем поступает в выносящие жаберные сосуды, впадающие в предсердия. Частично кровь из капилляров вен и артерий вытекает в мелкие лакуны, и потому кровеносную систему головоногих следует считать почти замкнутой. Кровь головоногих содержит дыхательный пигмент - гемоцианин, в состав которого входит медь, поэтому при окислении кровь голубеет.
Выделительная система представлена двумя или четырьмя (у Nautilus) почками. Внутренними концами они открываются в околосердечную сумку (перикард), а наружными - в мантийную полость. Продукты выделения поступают в почки из жаберных вен и из обширной перикардиальной полости. Дополнительно выделительную функцию выполняют перикардиальные железы, образованные стенкой, перикарда.
Половая система, размножение и развитие. Головоногие - раздельнополые животные. У некоторых видов хорошо выражен половой диморфизм, например у аргонавта (Argonauta). Самка аргонавта крупнее самца (рис. 243) и в период размножения выделяет вокруг тела при помощи особых желез на щупальцах тонкостенную пергаментоподобную выводковую камеру для вынашивания яиц, похожую на спиральную раковину. Самец аргонавта в несколько раз меньше самки и имеет особое удлиненное половое щупальце, заполняемое в период размножения половыми продуктами.
Гонады и половые протоки непарные. Исключение составляет наутилус, у которого сохранились парные протоки, отходящие от непарной гонады. У самцов семяпровод переходит в сперматофорную сумку, где сперматозоиды склеиваются в особые пакеты - сперматофоры. У каракатицы сперматофор имеет форму шашки; его полость заполнена сперматозоидами, а выходное отверстие закрыто сложной пробкой. В период размножения самец каракатицы при помощи полового щупальца с ложковидным концом передает сперматофор в мантийную полость самки.
330
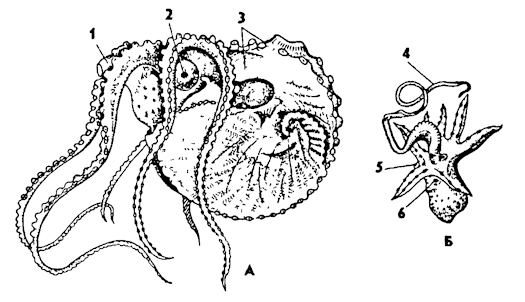
Рис. 243. Моллюск Аргонавт (Argonauta): А - самка, Б - самец; 1 - воронка, 2 - глаз, 3 - раковина, 4 - гектокотиль, 5 - воронка, 6 - глаз (по Догелю)
Там пробка разбухает и взрывается. Спермии входят в мантийную полость, где и происходит оплодотворение яиц. У самца аргонавта во время брачного поведения половое щупальце автоматически отрывается (аутотомия), плывет направленно и активно заползает в мантийную полость самки. Сперматофоры выходят из полости полового щупальца, раскрываются и спермин оплодотворяют яйца. Зоолог XIX в. Жорж Кювье обнаружил в мантийной полости самок аргонавтов половое щупальце с присосками, принял его за новый вид паразита, относящегося к плоским червям, и назвал его Hectocotylus. Это название - гектокотиль - сохранилось за половым щупальцем головоногих. У самок головоногих имеются особые нидаментальные железы, впадающие в яйцевод и выделяющие оболочку вокруг яиц. Оплодотворение у головоногих наружно-внутреннее и происходит не в половых путях самки, а в ее мантийной полости.
Головоногие откладывают яйца обычно на дне. У некоторых видов наблюдается забота о потомстве. Так, самка аргонавта вынашивает яйца в выводковой камере, а осьминоги охраняют кладку яиц, которую размещают в укрытиях из камней или в пещерах. Развитие прямое, без метаморфоза. Из яиц выходят маленькие, вполне сформированные головоногие моллюски.
Современные головоногие относятся к двум подклассам: подкласс Наутилиды (Nautiloidea) и подкласс Колеоидеи (Coleoidea). К вымершим подклассам относятся: подкласс Аммониты (Ammonoidea), подкласс Бактриты (Bactritoidea) и подкласс Белемниты (Belemnoidea).
331
Подкласс Наутилиды (Nautiloidea)
Современные наутилиды включают один отряд Nautilida. Он представлен лишь одним родом Nautilus, к которому относится всего несколько видов. Ареал распространения Nautilus ограничен тропическими областями Индийского и Тихого океанов. Ископаемых наутилид насчитывается более 2500 видов. Это древняя группа головоногих, известная с кембрия.
Наутилиды обладают многими примитивными особенностями: наличием наружной многокамерной раковины, несросшейся воронкой, многочисленными щупальцами без присосок, проявлением метамерии (четыре ктенидия, четыре почки, четыре предсердия). Сходство наутилид с низшими раковинными моллюсками проявляется в строении нервной системы из тяжей без обособленных ганглиев, а также в строении целомодуктов.
Наутилус относится к бентопелагическим головоногим. Он плавает в толще воды "реактивным" способом, выталкивая воду из воронки. Многокамерная раковина обеспечивает плавучесть его тела и опускание на дно. Наутилус издавна был объектом промысла из-за красивой перламутровой раковины. Из раковин наутилуса изготовлено много изысканных ювелирных изделий.
Подкласс Колеоидеи (Coleoidea)
Coleoidea в переводе с латинского означает "жесткие". Это жесткокожие моллюски, лишенные раковины. Колеоидеи - процветающая группа современных головоногих, включает четыре отряда, к которым относится около 650 видов.
Общими особенностями подкласса являются: отсутствие развитой раковины, сросшаяся воронка, щупальца с присосками.
В отличие от наутилид у них только два ктенидия, две почки и два предсердия. Coleoidea обладают высоким развитием нервной системы и органов чувств. Наибольшим числом видов характеризуются следующие три отряда.
Отряд Каракатицы (Sepiida). Наиболее характерными представителями отряда являются каракатицы (Sepia) и спирула (Spirula) с рудиментами внутренней раковины. У них 10 щупалец, два из которых ловчие. Это нектобентосные животные, держатся у дна и способны активно плавать.
Отряд Кальмары (Teuthida). Сюда относятся многие промысловые кальмары: Todarodes, Loligo и др. У кальмаров иногда сохраняется рудимент
332
раковины в виде ровогой пластинки под кожей на спине. У них 10 щупалец, как и у предыдущего отряда. Это в основном нектонные животные, активно плавающие в толще воды, имеющие торпедовидную форму тела (рис. 244).
Отряд Восьминогие (Octopoda). Это эволюционно продвинутая группа головоногих без следов раковины. У них восемь щупалец. Выражен половой диморфизм. У самцов развивается половое щупальце - гектокотиль. Сюда относятся разнообразные осьминоги (рис. 245). Большинство осьминогов ведет придонный образ жизни. Но среди них имеются нектонные и даже планктонные формы. К отряду Octopoda относится род Argonauta - аргонавт, у которого самка выделяет особую выводковую камеру.
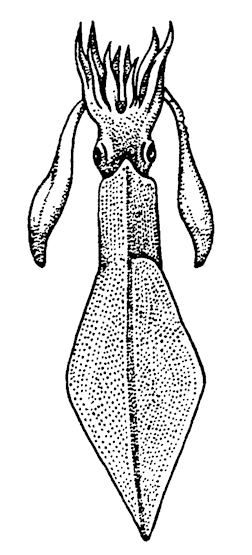
Рис. 244. Кальмар Loligo (из Догеля)
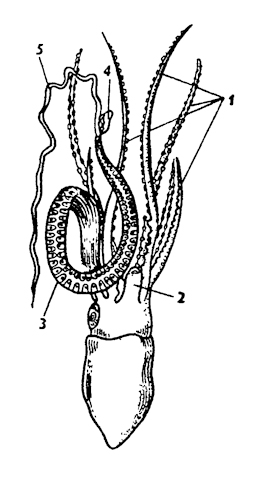
Рис. 245. Осьминог (самец) Ocythoe (по Пельзнеру): 1 - щупальца, 2 - воронка, 3 - гектокотиль, 4 - мешочек, 5 - концевая нить
333
Практическое значение головоногих
Головоногие моллюски - промысловые животные. Мясо каракатиц, кальмаров и осьминогов используется в пищу. Мировой улов головоногих в настоящее время достигает более 1600 тыс.т. в год. Каракатиц и некоторых осьминогов добывают также с целью получения чернильной жидкости, из которой изготавливают натуральную тушь и чернила высшего качества.
Палеонтология и филогения головоногих
Самой древней группой головоногих считают наутилид, ископаемые раковины которых известны уже по кембрийским отложениями. Примитивные наутилиды имели невысокую коническую раковину всего лишь с несколькими камерами и с широким сифоном. Предполагается, что головоногие произошли от древних ползающих раковинных моллюсков с простой конической раковиной и плоской подошвой, как у некоторых ископаемых моноплакофор. По-видимому, существенный ароморфоз в возникновении головоногих заключался в появлении первых перегородок и камер в раковине, что положило начало развитию у них гидростатического аппарата и определило возможность всплывать, отрываясь от дна. По-видимому, параллельно происходило формирование воронки и щупалец. Раковины древних наутилид были разнообразными по форме: длинные конические и плоские спирально закрученные с разным числом камер. Среди них встречались и гиганты до 4-5 м (Endoceras), которые вели придонный образ жизни. Наутилиды претерпели в процессе исторического развития несколько периодов расцвета и угасания и просуществовали до наших дней, хотя и представлены сейчас всего лишь одним родом Nautilus.
В девоне параллельно с наутилидами начинает встречаться особая группа головоногих - бактриты (Bactritoidea), меньшие по размерам и менее специализированные, чем наутилиды. Предполагается, что эта группа головоногих произошла от общих пока неизвестных предков с наутилидами. Бактриты оказались эволюционно перспективной группой. Они дали начало двум ветвям развития головоногих: аммонитам и белемнитам.
Подкласс аммонитов (Ammonoidea) появился в девоне и вымер в конце мела. В период расцвета аммониты успешно конкурировали с наутилидами, численность которых в это время заметно падала. Нам трудно судить о преимуществах внутренней организации аммонитов только по ископаемым раковинам. Но раковина аммонитов была более совершенной,
334
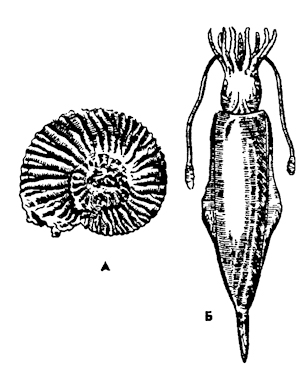
Рис. 246. Ископаемые головоногие: А - аммонит, Б - белемнит
чем у наутилид: более легкой и прочной. Перегородки между камерами у аммонитов были не гладкими, а волнистыми, а линии перегородок на раковине зигзагообразными, что увеличивало прочность раковины. Раковины аммонитов были спирально закрученными. Чаще обороты спирали раковин аммонитов располагались в одной плоскости, а реже имели форму турбоспирали (рис. 246, А). По некоторым отпечаткам тела ископаемых остатков аммонитов можно предполагать, что у них имелось до 10 щупалец, возможно, были два ктенидия, клювообразные челюсти, чернильный мешок. Это свидетельствует о том, что у аммонитов, по-видимому, произошла олигомеризация метамерных органов. По данным палеонтологии, аммониты были экологически более разнообразными, чем наутилиды, и среди них встречались нектонные, бентосные и планктонные формы. Большинство аммонитов имели небольшие размеры, но встречались и гиганты с диаметром раковины до 2 м. Аммониты были одними из самых многочисленных морских животных в мезозое, а их ископаемые раковины служат руководящими формами в геологии для определения возраста пластов.
Другая ветвь эволюции головоногих, гипотетически выводимая от бактритов, была представлена подклассом белемнитов (Belemnoidea). Белемниты появились в триасе, процветали в меловом периоде и вымерли в начале кайнозойской эры. По своему внешнему облику они уже ближе к современному подклассу Coleoidea. По форме тела они напоминают современных кальмаров (рис. 246, Б). Однако белемниты существенно отличались от них наличием тяжелой раковины, которая обрастала мантией. Раковина белемнитов была коническая, многокамерная, покрытая кожей. В геологических отложениях сохранились остатки раковин и особенно их концевые пальцеобразные рострумы, которые образно названы "чертовыми пальцами". Белемниты нередко были очень крупными: их длина достигала нескольких метров. Вымирание аммонитов и белемнитов, вероятно, было связано с усилившейся конкуренцией с костистыми рыбами. И вот в кайнозое на арену жизни выходит новая группа головоногих - колеоидеи (подкласс Coleoidea), лишенные раковин, с быстрым реактивным движением, со сложноразвитой нервной системой и органами чувств. Они-то и стали "приматами" моря и могли на равных конкурировать как хищники с рыбами. Эта группа головоногих появилась еще
335
в мелу, но высшего расцвета достигла в кайнозойскую эру. Есть основание считать, что Coleoidea имеют общие корни происхождения с белемнитами.
Экологическая радиация головоногих. Экологическая радиация головоногих представлена на рисунке 247. От примитивных раковинных бентопелагических форм, способных всплывать благодаря гидростатическому аппарату, определилось несколько путей экологической специализации. Наиболее древние экологические направления были связаны с радиацией наутилид и аммонитов, которые плавали на разных глубинах и образовывали специализированные раковинные формы бентопелагических головоногих. От бентопелагических форм прослеживается переход к бентонектонным (типа белемнитов). У них раковина становится внутренней, и ее функция плавательного аппарата ослабевает. Взамен у них развивается главный движитель - воронка. Позднее они дали начало безраковинным формам. Последние претерпевают бурную экологическую радиацию, образовав нектобентосные, нектонные, бентосные и планктонные формы.
Главными представителями нектона являются кальмары, но есть и быстроплавающие с узким торпедовидным телом осьминоги, каракатицы. К составу нектобентоса в основном относятся каракатицы, часто плавающие
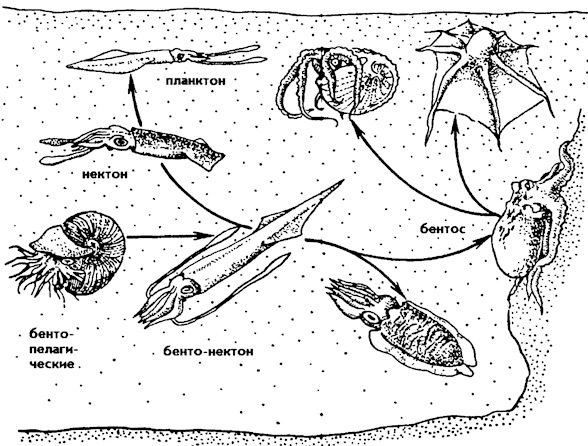
Рис. 247. Экологическая радиация головоногих
336
или лежащие на дне, к бентонектону - осьминоги, которые больше ползают по дну, чем плавают. К планктону относятся зонтикообразные, или студенистые, осьминоги, палочковидные кальмары.
337
3W.SU ©® 2015