Зоология беcпозвоночных
Этапы филогении животного мира
Современная система животного мира представляет собой результат эволюции и сопоставима с конечными разветвлениями общего родословного "древа". Нижние ветви "древа" представлены более древними по происхождению группами, сохранившими наиболее примитивные черты организации, а верхние ветви обладают более прогрессивными особенностями. Однако следует иметь в виду, что каждый современный таксон обладает как древними признаками (плезиоморфными), общими с предками, так и новыми особенностями, отличающими их от последних (апоморфными). Анализ современной системы, базирующейся на сравнительно-анатомических, эмбриологических, генетических, палеонтологических данных, позволяет косвенно судить о филогенетических связях таксонов и этапности эволюционного процесса. Данные палеонтологической летописи свидетельствуют о том, что большинство современных типов животных существовали уже в кембрии или силуре. Поэтому происхождение типов не имеет палентологических подтверждений и базируется исключительно на косвенных доказательствах.
Рассмотрим, как выглядят филогенетические построения для животного мира на современном этапе развития зоологии. По схеме филогенетического древа животных (рис. 405) проследим основные этапы в ходе эволюционного процесса. Детально не будем останавливаться на различных гипотезах и их доказательствах о происхождении различных систематических групп животных, так как они рассмотрены в предшествующих главах.
У основания филогенетического "древа" животных находятся одноклеточные (подцарство Protozoa). Это самый примитивный уровень организации животных, так как все функции одноклеточного организма выполняются одной клеткой и ее специализированными структурами - органеллами. В пределах подцарства наблюдаются переходы к многоклеточности (полиэнергидность, колониальность). Предполагается, что древние незеленые колонии жгутиконосцев осуществили исторический переход к многоклеточности у животных.
Подцарство многоклеточных (Metazoa) представляет собой качественно более высокий уровень организации животных, у которых различные функции организма выполняются специализированными клетками или многоклеточными органами. Среди современных многоклеточных можно выделить несколько уровней организации, которые отражают этапность эволюционного процесса многоклеточных.
К самым примитивным Metazoa относятся фагоцителлообразные (надраздел Phagocytellozoa), включающие один тип пластинчатых (Placozoa). По своей организации Placozoa представляют живую модель гипотетического
572
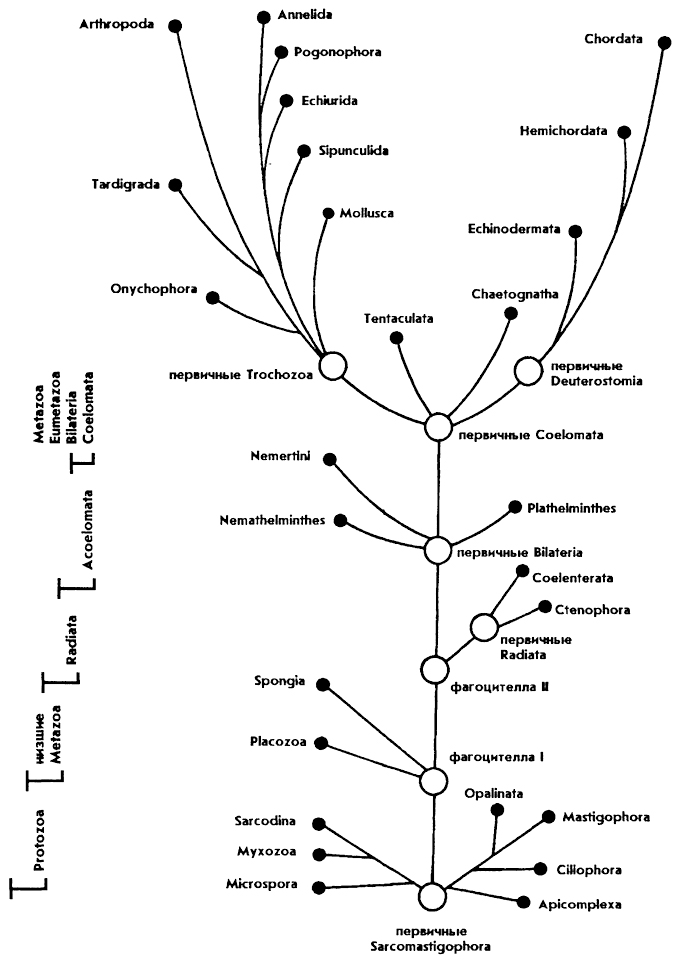
Рис. 405. Схема филогении животного мира
573
предка многоклеточных - фагоцителлы (по Мечникову). Им свойственны только два основных типа клеток: двигательные - жгутиковые (кинетобласт) и пищеварительные - амебоидные (фагоцитобласт). Такая функциональная дифференциация клеток в организме существенно отличает многоклеточных от колониальных одноклеточных с однородными клетками.
Следующий, более продвинутый уровень организации многоклеточных представляют паразои (надраздел Parazoa), к которым из современных животных относятся только губки (тип Spongia). В отличие от фагоцителлообразных губки состоят из множества специализированных клеток, выполняющих разнообразные функции (покровную, скелетную, пищеварительную и др.). Однако у губок еще отсутствуют оформленные ткани, органы, нет нервных и чувствующих клеток. Низкий уровень организации губок проявляется в их высокой регенерационной способности, взаимопревращаемости клеток и экскурвации пластов клеток в процессе онтогенеза.
Согласно современной гипотезе А. В. Иванова, эти низшие надразделы многоклеточных (Phagocytellozoa, Parazoa) произошли от гипотетического предка - фагоцителлы. Их ранние фазы развития: бродяжка у трихоплаксов (тип Placozoa) и личинка-паренхимула у губок (тип Spongia) - модель фагоцителлы. При этом Placozoa сохранили план строения фагоцителлы, а у Spongia двигательные клетки (кинобласт) переместились внутрь тела и стали выполнять функцию водно-двигательной системы, обеспечивающей фильтрацию.
Высшие многоклеточные образуют надраздел Eumetazoa. Их тело состоит уже из оформленных тканей, органов, выполняющих разные функции. Впервые на этом этапе развития животных появляются нервные и чувствующие клетки. Среди Eumetazoa также можно выделить несколько групп организмов, находящихся на разных уровнях организации, отражающих эволюционную тенденцию их прогрессивного развития.
Низшую ступень среди них образуют двуслойные животные с радиальной симметрией (раздел Radiata, или Diploblastica), к которым относятся типы кишечнополостных (Coelenterata) и гребневиков (Ctenophora), ведущих малоактивный или прикрепленный образ жизни. Их тело формируется из двух зародышевых листков: наружного - эктодермы и внутреннего - энтодермы. Согласно современным воззрениям, Radiata, как и низшие надразделы многоклеточных произошли от фагоцителлы. Об этом свидетельствует строение личинок кишечнополостных, похожих на паренхимулу губок. При эволюционном переходе от фагоцителлы к кишечнополостным кинобласт дал начало эктодерме, и у подвижных форм (личинки кишечнополостных и гребневики) сохранилась ее первоначальная двигательная функция. Coelenterata перешли в основном
574
к прикрепленному образу жизни. Оба типа сохранили первичную радиальную симметрию.
Высшую ступень в развитии Eumetazoa представляют билатеральные трехслойные животные (раздел Bilateria), ведущие, как правило, активный образ жизни. У них различают передний и задний концы тела. Рот смещен на брюшную сторону тела, что привело к формированию билатеральной симметрии. Однако целый ряд групп из Bilateria вторично вернулись к радиальной симметрии (иглокожие), сохранив частично билатеральность строения. Тело Bilateria образуется из трех зародышевых пластов: эктодермы, энтодермы и промежуточного слоя - мезодермы.
Ранее считали, что Bilateria произошли от Radiata. Теперь же имеются доказательства их независимого происхождения от фагоцителлоподобных предков. Выяснилось, что у Bilateria и Radiata нет симплезиоморфности по ряду важнейших признаков, присущих предкам. Так, паренхиматозность строения сохраняют низшие Bilateria (типы Plathelminthes и частично Nemathelminthes), а двуслойность тканевого строения, функционально близкая к двум основным типам клеток фагоцителлообразных (кинобласт и фагоцитобласт), наблюдается только у Radiata. О самостоятельности этих двух путей эволюционного развития свидетельствуют также существенные различия их апоморфных (эволюционно продвинутых) признаков. Так, у Radiata изначально образовалась кишечная полость, достигшая высокой сложности строения, развивались разные формы радиальной симметрии, особые опорные структуры в мезоглее. А у Bilateria кишечник возник позднее из внутреннего слоя паренхимы. Большое прогрессивное значение у них имело возникновение третьего зародышевого листка - мезодермы и формирование из него мускулатуры, гонад. Впервые у Bilateria появляется выделительная система органов (протонефридии). Кроме того, с развитием билатеральной симметрии, связанной с поступательными движениями Bilateria, обособляется головной отдел тела с нервными узлами и органами чувств.
Таким образом, надраздел Eumetazoa развивался в двух направлениях (Radiata и Bilateria) от исходных фагоцителлообразных предков.
В эволюции Bilateria различают два этапа: образование низших бесполостных (подраздел Acoelomata) и высших целомических животных (подраздел Coelomata).
Бесполостные (Acoelomata), или низшие черви (Scolecida), объединяют группу типов, у которых отсутствует вторичная полость тела - целом. У низших групп этого подраздела - плоских червей (тип Plathelminthes) и некоторых круглых (тип Nemathelminthes) промежутки между органами заняты паренхимой, а у более продвинутых групп круглых червей и немертин (тип Nemertini) образуется первичная полость тела - схизоцель за счет разрушения паренхимы. Наиболее примитивными
575
среди современных Acoelomata принято считать бескишечных турбеллярий (Acoela), сохраняющих во многом черты фагоцителлообразных предков.
Высший подраздел билатеральных животных образуют целомические (Coelomata), у которых образуется вторичная полость тела - целом, выстланная целомическим эпителием мезодермального происхождения.
Образование целома завершило в эволюции животных формирование внутренней среды организма, что обеспечило гомеостаз и автономизацию жизненных процессов. Их эволюция связана с формированием кровеносной системы, метанефридиев, целомодуктов, развитой нервной системы и органов чувств.
Целомические животные дали широкий спектр таксонов, филогенетические отношения которых еще недостаточно расшифрованы. Однако сравнительное морфологическое изучение наиболее примитивных Coelomata из различных типов и их онтогенеза позволило выявить у них некоторые общие плезиоморфные признаки, дающие представление об уровне организации их общих предков.
Вероятно, что первичные Coelomata обладали следующими плезиоморфными признаками: аметамерией, сочетанием развитой первичной полости тела (схизоцеля) и формирующегося целома, отсутствием кровеносной системы, наличием лестничной нервной системы, близкой к ортогону, но с выраженным окологлоточным кольцом. Роль органов выделения могли выполнять протонефридии или целомодукты. Покровы, по-видимому, включали поля ресничного эпителия, а мускулатура была представлена кольцевыми и продольными мышцами. Все указанные особенности указывают на родство и возможное происхождение Coelomata от турбелляриеподобных предков.
Проследим основные пути дальнейшей эволюции Coelomata. Ранее считали, что таких направлений было два: к первичноротым (надтип Protostomia) и вторичноротым (надтип Deuterostomia), отличавшихся типом закладки мезодермы и формированием рта в эмбриогенезе. Новейшие данные по эмбриологии первичноротых показали, что у них наблюдается большое разнообразие в закладке целома и кишечника и при этом нередко проявляется параллелизм со вторичноротыми.
В настоящее время выделяют пять основных направлений в эволюции Coelomata, которые дали начало следующим надтипам (по А. В. Иванову): трохофорным (Trochozoa), вторичноротым (Deuterostomia) и занимающим промежуточное положение погонофорам (Pogonophora), щупальцевым (Tentaculata) и щетинкочелюстным (Chaetognatha).
Трохофорные (Trochozoa) отличаются от остальных надтипов особым типом спирального дробления яйца, детерминированностью эмбриогенеза,
576
формированием у большинства типов личинки трохофоры или близкой к ней фазы развития. У многих, за некоторыми исключениями, рот первичен, т.е. соответствует бластопору. Закладка мезодермы у трохофорных может быть телобластической, а нередко и энтероцельной.
Первичные трохофорные были аметамерными и от них, по-видимому, произошли Echiurida, Sipunculida с цельным целомом.
Ведущей эволюционной тенденцией в надтипе трохофорных явилось развитие метамерии, вначале внешней, а затем и внутренней. Тип Mollusca отделился от общего ствола трохофорных довольно рано, по-видимому, от олигомерных предков, а возможно, и от аметамерных. Решение этой проблемы связано с выяснением природы метамерии у низших групп моллюсков. Примерно на этом же этапе шло формирование типов Onychophora и Tardigrada с неметамерной полостью и лишь с наружной сегментацией. Узловым моментом в эволюции трохофорных было образование предковой группы первичных кольчецов, от которых произошли полимерные кольчецы Annelida с метамерным целомом и олигомерные - членистоногие Arthropoda, у которых сформировалась смешанная полость тела - миксоцель и наружный хитиновый скелет.
Надтип вторичноротые (Deuterostomia) образует вторую крупную ветвь в эволюции целомических животных. К ним относятся типы: тип Иглокожие (Echinodermata), тип Полухордовые (Hemichordata) и тип Хордовые (Chordata). Тело вторичноротых в эмбриогенезе формируется из трех сегментов; рот у них закладывается вторично, мезодерма образуется энтероцельным путем, кожа состоит из эктодермального эпителия и мезодермального кутиса. Вершиной эволюции надтипа Deuterostomia явился тип Chordata. Среди хордовых наивысшего прогресса достигли позвоночные животные (подтип Vertebrata) с развитой нервной системой, органами чувств, сложным поведением. Высшие позвоночные животные - птицы и млекопитающие в процессе эволюции достигли теплокровности, что позволило им занять особенно широкий спектр экологических ниш на планете.
Промежуточное положение между трохофорными и вторичноротыми животными занимают надтипы: Chaetognatha и Tentaculata. У них имеются признаки сходства как с трохофорными, так и со вторичноротыми животными, а также имеются особые уникальные плезиоморфные признаки, общие с древними предковыми Coelomata.
Спорным остается вопрос о происхождении типа погонофор. Согласно одной точке зрения, погонофоры занимают также промежуточное положение между трохофорными и вторичноротыми животными, а согласно другой, - погонофоры близки по организации к кольчатым червям и их следует относить к трохофорным животным.
577
3W.SU ©® 2015