Зоология беcпозвоночных
Проблема происхождения многоклеточных
Вопрос о происхождении многоклеточных животных имеет большое теоретическое значение, так как представляет собой основу для понимания эволюции организации животных и их индивидуального развития.
Существует множество гипотез о происхождении многоклеточных, однако большинство ученых считает доказанным происхождение Metazoa от Protozoa. Все структурные компоненты клетки Protozoa частично или полностью идентичны таковым у Metazoa. Кроме того, в пределах Protozoa прослеживается тенденция перехода к многоклеточности. Это проявляется у полиэнергидных простейших с многочисленными ядрами (опалина, миксоспоридии, некоторые инфузории, радиолярии, фораминиферы) и у колониальных форм, например у вольвоксовых жгутиконосцев. В некоторых случаях у простейших наблюдается даже многоклеточность отдельных фаз развития, например спор у миксоспоридии.
Проявления полиэнергидности и колониальности у Protozoa послужили основанием к разработке гипотез о происхождении Metazoa. Гипотезы происхождения многоклеточных подразделяются на две группы - колониальные и полиэнергидные, в зависимости от того, какие группы простейших принимаются за исходные в эволюции.
I. Колониальные гипотезы происхождения Metazoa базируются на признании в качестве предков колониальных Protozoa (рис. 67, табл. 4).
1. Первую колониальную гипотезу происхождения Metazoa разработал зоолог-эволюционист Э. Геккель (1874), гипотеза получила название "гастреи" (рис. 67, I). Он считал, что протозойным предком Metazoa была "бластея" - шаровидная колония жгутиковых, похожая на стадию бластулы в развитии многих многоклеточных. В процессе эволюции от бластеи путем инвагинации (впячивания) могли возникнуть первые двуслойные многоклеточные с кишечной полостью, выстланной энтодермой. Этот гипотетический предок Metazoa был назван Э. Геккелем "гастреей" в связи со сходством со стадией гаструлы в развитии многоклеточных. Гастрея, по Геккелю, представляла плавающее двуслойное животное со ртом. Наружный слой жгутиковых клеток гастреи представлял эктодерму и выполнял двигательную функцию, а внутренний слой клеток (энтодерма) - пищеварительную. От гастреи, по его мнению, произошли прежде всего двуслойные животные - кишечнополостные.
2. Дальнейшее развитие теории "гастреи" продолжил О. Бючли (1884), предложивший ее новый вариант - гипотезу "плакулы" (рис. 67, II). Поправка Бючли к теоретическим рассуждениям Геккеля состояла в том, что он считал колониальных простейших типа "бластеи" эволюционно продвинутыми и предложил в качестве гипотетического колониального
100
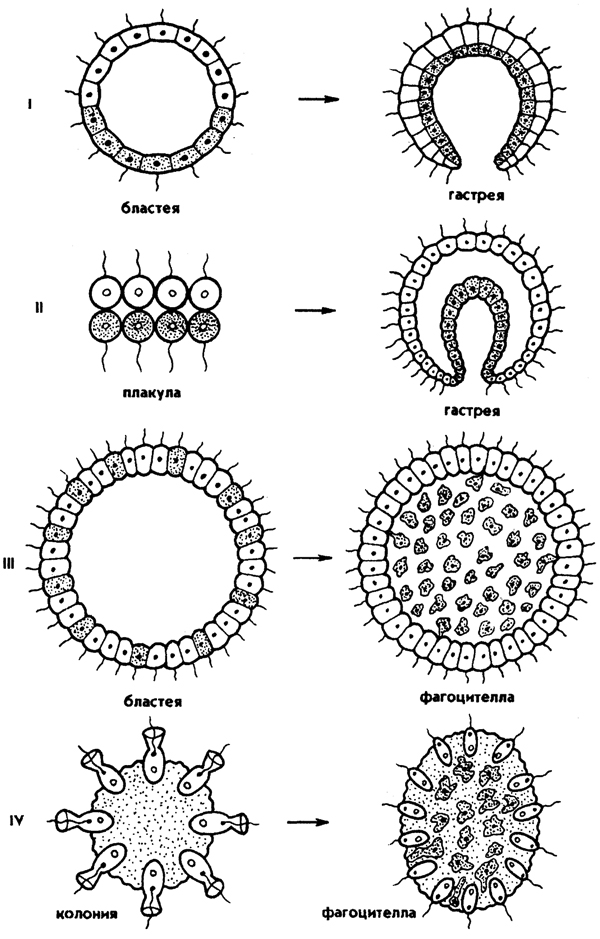
Рис. 67. Схема происхождения многоклеточных (колониальные): I - по гипотезе Геккеля, II - по гипотезе Бючли, III - по гипотезе Мечникова, IV - по гипотезе Иванова
101
Таблица 4. Колониальные гипотезы происхождения Metazoa
| Гипотеза | Колониальный предок из Protozoa | Процесс преобразования | Предок многоклеточных Metazoa | Живая модель предка |
| "Гастреи" (Геккель, 1874) | Бластея (однослойная шаровидная) | Инвагинация | Гастрея (двуслойная шаровидная со ртом) | Гаструла и личинка кишечнополостных |
| "Плакулы" (Бючли, 1884) | Плоская однослойная колония | Расслоение колонии и изгибание | Гастрея (двуслойная со ртом) | Трихоплакс, гаструла |
| "Фагоцителлы" (Мечников, 1882) | Бластея (однослойная шаровидная) | Иммиграция | Фагоцителла (из двух слоев без рта) | Личинка губок - паренхимула |
| "Фагоцителлы" (Иванов, 1967) | Колония воротничковых жгутиковых (без полости) | Иммиграция | Фагоцителла (из двух слоев без рта) | Паренхимула, трихоплакс |
предка более простую пластинчатую колонию одноклеточных типа современных Gonium. Путем расщепления такой пластинки на два слоя возник, по Бючли, гипотетический предок многоклеточных - "плакула". В дальнейшем из плакулы могла образоваться, по Бючли, гастрея путем чашевидного прогибания двуслойной пластинки. Натолкнуло Бючли на создание нового варианта гипотезы описание примитивного двуслойного многоклеточного животного - трихоплакса (Trichoplax), строение которого приближалось к плоским колониям жгутиковых. Автор предполагал, что подобные трихоплаксу животные могли быть промежуточными между "плакулой" и "гастреей". Тем более было обнаружено, что у Trichoplax нижний слой клеток способен к наружному пищеварению. Ползая, эти животные выделяют пищеварительные соки (ферменты), переваривающие бактериальную плёнку. В дальнейшей эволюции, по его мнению, из этого нижнего слоя клеток примитивных плакулоподобных организмов возникла энтодерма гастреи, а из верхнего - эктодерма.
102
3. Русский биолог И. И. Мечников в 1882 г. опубликовал другую колониальную гипотезу - "фагоцителлы", раскрывающую сущность происхождения многоклеточных. Гипотеза базировалась на обширных исследованиях автора. Мечников открыл явление фагоцитоза - внутриклеточного пищеварения у многоклеточных и считал этот способ переваривания пищи более примитивным, чем полостное пищеварение. По его мнению, первые многоклеточные были примитивнее "гастреи" по организации и не имели еще пищеварительной полости и полостного пищеварения. Для выяснения вопроса о гипотетическом предке Metazoa И. И. Мечников пристально изучал онтогенез примитивных многоклеточных - губок. Им было обнаружено, что образование двуслойной фазы развития у губок происходит не путем инвагинации бластулы, а путем иммиграции отдельных клеток наружного слоя в полость зародыша (бластоцель). Личинка губок с паренхимными клетками внутри была названа паренхимулой. И. И. Мечников рассматривал паренхимулу как прообраз или живую модель гипотетического предка многоклеточных - фагоцителлы (рис. 67, III). Это название предка связано со способом питания - фагоцитозом, который осуществлялся в паренхиматозных клетках. По его мнению, фагоцителла могла возникнуть из шаровидных колоний жгутиконосцев путем иммиграции части клеток внутрь колонии. При этом наружные клетки со жгутиками выполняли функцию движения (кинобласт), а внутренние - утрачивали жгутики, становились амебоидными и выполняли функцию фагоцитоза (фагоцитобласт).
Гипотеза фагоцителлы И. И. Мечникова завоевала широкое признание и нашла дальнейшее развитие в трудах многих ученых. Дополнения и существенные поправки к теории фагоцителлы внесли советские ученые А. А. Захваткин и А. В. Иванов. Так, А. А. Захваткин (1949) создал гипотезу синзооспоры. По его мнению, нельзя было принимать в качестве предка, многоклеточных колонии зеленых жгутиконосцев (типа Volvox), как это допускали предшественники, так как у вольвоксовых голофитный способ питания и зиготическая редукция хромосом, как у растений. Поэтому А. А. Захваткин предположил, что колониальные простейшие, давшие начало Metazoa, обладали не голофитным, а голозойным типом питания и имели гаметическую редукцию хромосом в онтогенезе. В связи с тем, что у всех Metazoa эмбриогенез протекает в пределах яйцевой оболочки и дробление зародыша вначале палинтомическое и только после выхода зародыша из яйца дробление становится монотомическим, Захваткин предположил, что колониальный предок в своем индивидуальном развитии также проходил подобные фазы дробления.
Другая важная поправка касалась облика первого многоклеточного животного. Захваткин считал, что фагоцителла И. И. Мечникова отражает
103
облик не взрослого предка многоклеточных, а лишь его личинки - синзооспоры. А взрослая фаза предка многоклеточных, по мнению Захваткина, представляла сидячую форму колониального типа, похожую на губок. Но гипотеза синзооспоры не получила широкого распространения, так как трудно было допустить, чтобы сидячие колониальные формы могли дать дальнейшую эволюцию всех Metazoa.
Крупный современный зоолог А. В. Иванов (1967) синтезировал современные идеи по проблеме происхождения многоклеточных. За основу он принял гипотезу фагоцителлы Мечникова. Однако он предложил считать в качестве колониального предка Metazoa колонию типа воротничковых жгутиковых, имеющих голозойный способ питания, что соответствовало взглядам А. А. Захваткина. Живой моделью фагоцителлы А. В. Иванов считает не столько личинку губок - паренхимулу, сколько трихоплакса, близкого по организации к фагоцителле. Фагоцителла, по А.В.Иванову, в процессе эволюции дала начало таким типам, как Губки (Spongia) и Пластинчатые (Placozoa), обладающим примитивным внутриклеточным пищеварением - фагоцитозом. Согласно взглядам А. В. Иванова, появление двуслойных животных со ртом, кишечной полостью и полостным пищеварением произошло значительно позднее, чем фагоцителлоподобных (рис. 68).
П. Полиэнергидные гипотезы происхождения многоклеточных исходят из того, что предками Metazoa были полиэнергидные простейшие.
Впервые идея происхождения Metazoa от полиэнергидных Protozoa была предложена Иерингом, а позднее активно защищалась югославским
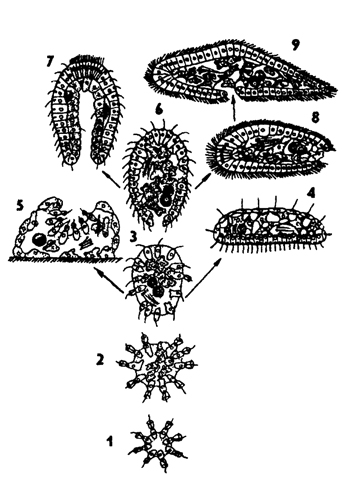
Рис. 68. Происхождение низших многоклеточных (по Иванову): 1 - колония воротничковых жгутиконосцев типа Sphaeroeca с монотомическим размножением, 2 - колония воротничковых жгутиконосцев типа Proterospongia с палинтомическим размножением и половым процессом, 3 - ранняя фагоцителла I без рта, 4 - пластинчатые (Placozoa) без рта, 5 - губки (Spongia) без рта и кишечника, 6 - поздняя фагоцителла II сортом, 7 - первичные кишечнополостные типа гастреи (двуслойные со ртом), 8 - первичные турбеллярии (тип Plathelminthes) - паренхиматозные со ртом, смещенным к брюшной стороне, трехслойные, 9 - бескишечные турбеллярии с дальнейшей дифференциацией клеток и смещением рта на брюшную сторону
104
зоологом Иованом Хаджи (1963). По мнению Хаджи, предками Metazoa были инфузории, а первыми многоклеточными - плоские черви (планарии). При этом процесс образования многоклеточности происходил путем целлюляризации, т. е. в клетке одноклеточного вокруг ядер обособлялись клетки. Например, по Хаджи, из эктоплазмы инфузорий и ядер - производных макронуклеуса возникали покровные клетки (эктодерма), из эндоплазмы и ядер - производных микронуклеуса образовывалась внутренняя паренхима (энтодерма и мезодерма), а из различных органелл - органы первого многоклеточного организма.
По названию процесса, якобы приведшего к образованию многоклеточности, полиэнергидную гипотезу часто называют еще и целлюлярной.
Идея о происхождении Metazoa от полиэнергидных Protozoa через целлюляризацию несомненно заслуживает внимания, однако конкретная аргументация гипотезы Хаджи сомнительна. Автором допущена некоторая механистичность в объяснении происхождения от Protozoa довольно высокоорганизованных групп трехслойных животных - червей. В этой гипотезе также не учтены: постулаты клеточной теории, теории зародышевых пластов, биогенетический закон, гомологичность структур, особенности жизненных циклов предков и потомков.
Обзор гипотез показывает, что в настоящее время наиболее серьезно аргументирована гипотеза фагоцителлы И. И. Мечникова, доработанная А. В. Ивановым с учетом всех достижений в этой области.
Классификация многоклеточных. Подцарство Metazoa в настоящее время подразделяют на три надраздела с разным уровнем организации: надраздел Фагоцителлообразные (Phagocytellozoa), примитивные многоклеточные, надраздел Паразои (Parazoa) и высшие, или собственно многоклеточные, надраздел Эуметазои (Eumetazoa).
Phagocytellozoa обладают лишь двумя основными типами клеток: двигательными со жгутиками (кинобласт) и фагоцитарными, или пищеварительными (фагоцитобласт). Клетки легко взаимопревращаются. Пищеварение внутриклеточное. Parazoa состоит из множества типов клеток, но у этих животных отсутствуют ткани, органы. Пищеварение внутриклеточное. Eumetazoa обладают широкой дифференциацией клеток, тканей, органов. Пищеварение может быть как внутриклеточное, так и полостное. Ниже следует обзор надразделов многоклеточных.
105
3W.SU ©® 2015