Зоология беcпозвоночных
Класс Насекомые открыточелюстные (Insecta-Ectognatha)
Насекомые открыточелюстные - основной класс шестиногих (Hexapoda). Как и у всех шестиногих, тело насекомых подразделяется на голову с усиками и тремя парами ротовых конечностей, трехсегментную грудь с тремя парами ног и брюшко, лишенное развитых конечностей. Дыхание трахейное.
В отличие от класса Insecta-Entognatha, у насекомых ротовой аппарат открытый, отсюда название класса - Insecta-Ectognatha. У большинства насекомых имеются развитые крылья на средне- и заднегруди и сложные фасеточные глаза на голове. Рудименты брюшных ножек модифицированы в половые придатки на заднем конце тела. Но у низших насекомых могут присутствовать и грифельки - рудименты брюшных ножек.
Насекомые достигли высокого прогресса среди трахейнодышащих и всех членистоногих. Это связано с их высокоразвитыми адаптациями к жизни на суше. По сравнению с многоножками и скрыточелюстными насекомые претерпели широкую экологическую радиацию, захватив самые разнообразные экологические ниши на суше. Некоторые из них вторично перешли в воду и играют существенную роль в пресноводных биоценозах.
Решающее значение в освоении суши, насекомыми сыграли их адаптации в строении покровов, морфофункциональных особенностях выделительной, трахейной систем, в совершенствовании и функциональном многообразии органов движения - ног, ротовых аппаратов и появлении органов полета - крыльев. Кроме того, особенности эмбрионального и постэмбрионального развития насекомых обеспечили им большую независимость от факторов сухопутной среды обитания. В отличие от прочих трахейнодышащих, насекомые смогли значительно расширить свои экологические возможноспд.
Всего известно около 1 млн. видов насекомых. Однако эта цифра, по-видимому, занижена. Некоторые энтомологи считают, что число видов насекомых достигает 2 и даже 3 млн. Ежегодно описывают десятки и
429
даже сотни новых видов насекомых. И потому очевидно, что классификация насекомых еще не завершена.
Это самая процветающая группа животных на Земле как по числу видов и особей, так и по широте пространственного распределения и экологической дифференциации.
Внешнее строение. Тело насекомых подразделено на три отдела: голову, грудь и брюшко (рис. 316). Размеры тела варьируют от 1-2 мм до 15-30 см.
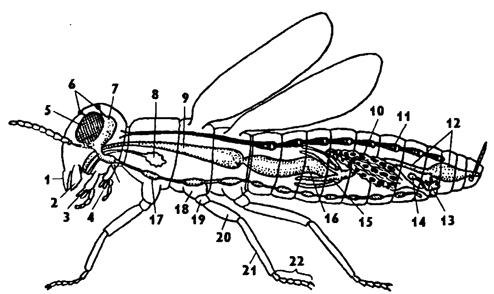
Рис. 316. Схема организации крылатого насекомого (по Хадорну): 1 - верхняя губа, 2 - мандибулы, 3 - максиллы, 4 - нижняя губа, 5 - глаза фасеточные, 6 - глазки простые, 7- мозг, 8 - слюнная железа, 9 - зоб, 10 - яичник, 11 - сердце, 12 - задняя кишка, 13 - придаточные железы, 14 - семяприемник, 15 - мальпигиевы сосуды, 16 - средняя кишка, 17 - брюшная нервная цепочка, 18 - тазик, 19 - вертлуг, 20 - бедро, 21 - голень, 22 - лапка
Голова слитная и состоит из акрона и четырех сегментов. Однако в последнее время принято считать, что акрон насекомых - сложное образование, состоящее из собственно предротовой лопасти и двух сегментов (антеннального и губного). В этом случае можно считать, что голова насекомого состоит из головной лопасти и шести сегментов.
На голове находятся антенны - придатки акрона и три пары челюстей: мандибулы - конечности второго головного сегмента, максиллы - конечности третьего сегмента и непарная нижняя губа, образованная в результате слияния второй пары максилл - конечностей четвертого сегмента. Первый головной сегмент интеркалярный, или вставочный, слабо развит и лишен конечностей.
На голове взрослых насекомых расположены сложные фасеточные глаза и 1-3 простых глазка. Три пары челюстей насекомых образуют ротовой аппарат открытого типа. В состав ротового аппарата входит также верхняя губа, представляющая подвижную пластинку на переднем крае головы. Верхняя губа прикрывает челюсти сверху. У некоторых насекомых в состав ротового аппарата может входить еще непарный вырост глотки - гипофаринкс, или "язычок".
Усики (антенны) - членистые придатки, отходящие от лицевой поверхности головы, обычно между глаз. Усики - органы осязания и обоняния.
430
Они могут быть нитевидными, четковидными, гребневидными, булавовидными, перистыми и др.
Ротовые части насекомых, представленные тремя парами конечностей, также весьма разнообразны. Различают грызущий, грызуще-лижущий, колюще-сосущий, сосущий и лижущий ротовые аппараты.
Первичным типом ротового аппарата у насекомых, по-видимому, следует считать грызущий (рис. 317). Подобный тип ротового аппарата свойствен низшим трахейным: многоножкам и низшим группам шестиногих. От грызущего типа ротового аппарата с наиболее полным составом ротовых частей прослеживаются морфологические переходы к другим специализированным типам ротовых аппаратов. Существуют и онтогенетические доказательства примитивности грызущего ротового аппарата, который наблюдается у личинок всех насекомых, развивающихся с полным метаморфозом. Так, у многих насекомых со специализированным ротовым аппаратом (бабочек, мух, пчел) личинки имеют грызущие ротовые части.
Грызущий ротовой аппарат свойствен тараканам, прямокрылым, жукам и другим насекомым (рис. 317, А). В состав грызущего аппарата входят: верхние челюсти - мандибулы жующего типа с зубцами на внутренней поверхности, максиллы, состоящие из двух базальных члеников (кардо), стволика (стипеса), челюстных щупиков и двух жевательных лопастей (наружной и внутренней), и нижняя губа, состоящая из сросшейся второй пары нижних челюстей. Базальная пластинка нижней губы - подподбородок соответствует кардо максилл. Причлененный к подподбородку подбородок гомологичен стипесу максилл. Нижнегубные щупики соответствуют щупикам нижних челюстей, а две пары язычков - жевательным лопастям максилл. Сверху грызущий ротовой аппарат прикрыт верхней губой.
Остальные типы ротовых аппаратов насекомых - производные от грызущего. Это связано с переходом к питанию жидкой пищей.
Среди специализированных ротовых аппаратов насекомых наименее измененным по сравнению с грызущим является грызуще-лижущий, или лакающий, ротовой аппарат пчелиных (пчелы, шмели). Ротовой аппарат пчелиных приспособлен для высасывания нектара из цветков. Эту функцию выполняет длинный хоботок, состоящий из вытянутых максилл и нижней губы. Жевательные лопасти максилл превращены в вытянутые заостренные к вершине лопаточки, а нижнечелюстные щупики радуцированы. Основную функцию слизывания нектара выполняет длинный язычок, образованный внутренними парными язычками нижней губы. Нижнегубые щупики длинные, прилегающие к язычку. Хоботок действует по типу капиллярной системы. Верхние челюсти пчелиных грызущего типа. При помощи мандибул пчелы строят соты и разжевывают пальцу.
431
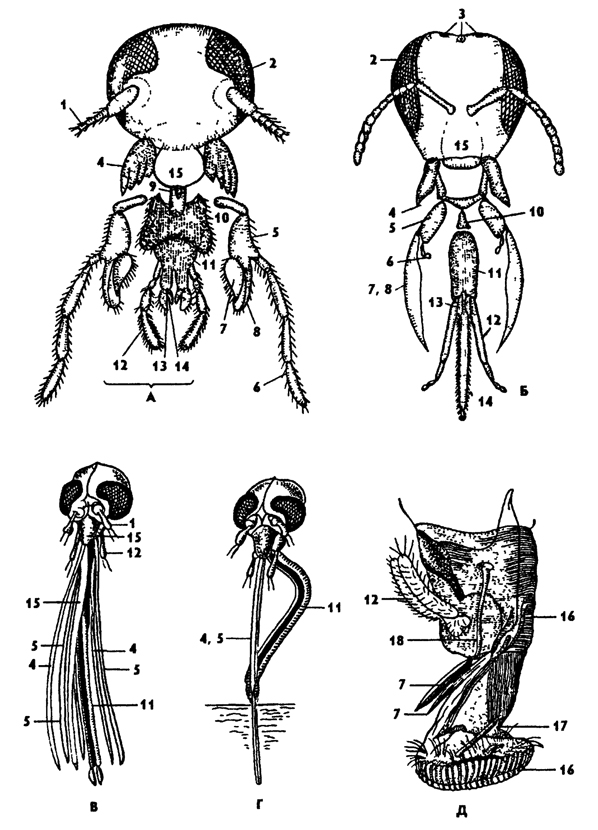
Рис. 317. Ротовые аппараты насекомых (из Хадорна): А - грызущий у таракана, Б - грызуще-лижущий у пчелы, В, Г - колюще-сосущий у комара, Д - лижущий у мухи; 1 - антенны, 2 - глаза сложные, 3 - глазки простые, 4 - мандибулы, 5 - стволик максилл,6 - челюстной щупик 7, 8 - нижние челюсти, жевательные лопасти, 9 - сочленение, 10 - подподбородок, 11 - подбородок нижней губы, 12 - губной щупик, 13, 14 - язычок, 15 - верхняя губа, 16 - нижняя губа, 17 - сочленение с лопастями, 18 - слюнные протоки
432
Более специализирован ротовой аппарат кровососущих комаров - колюще-сосущий (рис 317, В, Г). В состав колюще-сосущего хоботка входят все элементы ротового аппарата. Колющую часть хоботка составляют пять длинных иголочек (мандибулы, максиллы и гипофаринкс). Иглы вкладываются в желоб вытянутой нижней губы. Сверху желоб прикрыт длинной верхней губой, которая срослась в тонкую трубочку. При укусе комар прокалывает кожу жертвы иглами хоботка, а по трубочке верхней губы как по капилляру кровь поднимается к ротовому отверстию. Нижняя губа при этом коленчато изгибается, упирается в субстрат и служит опорой колющей части хоботка, погруженной в покровы (рис 317, Г).
У клопов ротовой аппарат сходного типа. Колющую часть аппарата составляют верхние и нижние челюсти, а нижняя губа членистая с желобом, в который вкладываются, как в ножны, челюсти.
Сосущий ротовой аппарат характерен для бабочек, высасывающих нектар из цветков с глубокими нектарниками (рис 318). Это один из самых специализированных ротовых аппаратов у насекомых. От всего состава ротовых частей здесь сохранились лишь нижние челюсти, образующие длинный спирально закрученный хоботок. При этом мандибулы редуцировались, а нижняя губа с опущенными щупиками превратилась в органы обоняния.
Лижущий ротовой аппарат мух относится к числу самых специализированных аппаратов насекомых (рис 317, Д). У мух имеется хоботок, при помощи которого они слизывают жидкую пищу, в том числе и нектар. Хоботок образован нижней губой с фильтрующими лопастями на конце. Муха способна высасывать жидкость или отфильтровывать ее из смеси с твердыми частицами. На хоботке имеется желобок, прикрытый верхней губой. В желобке расположен язычок - гипофаринкс, способствующий сосанию. Остальные ротовые части рудиментарны. У хищных и кровососущих мух помимо лижущей губы имеются режущие челюсти.
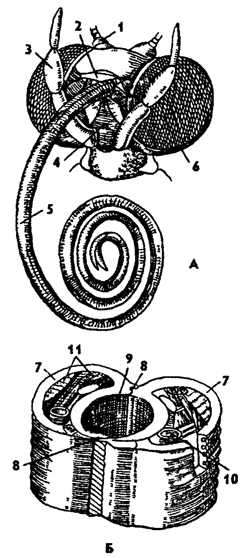
Рис. 318. Сосущий ротовой аппарат бабочки (по Веберу): А - голова бабочки с хоботком, Б - участок хоботка на поперечном разрезе ); 1 - основание усиков, 2 - верхняя губа, 3 - нижнегубной щупик, 4 - нижняя губа, 5 - хоботок - нижние челюсти, 6 - глаз, 7 - нижние челюсти, 8 - соединение челюстей, 9 - полость хоботка, 10 - трахеи, 11 - мускулатура хоботка
433
У некоторых насекомых ротовой аппарат редуцирован и не функционирует, как, например, у имаго поденок. Взрослые стадии поденок живут всего несколько часов и выполняют лишь функцию размножения, после чего отмирают. Питание поденок происходит только на личиночной фазе развития.
Рассмотренные типы ротовых аппаратов свидетельствуют о широкой экологической радиации насекомых, а также и о конвергенции в строении ротовых аппаратов в связи со сходством в способах питания у некоторых групп насекомых. Так, у всех насекомых, питающихся жидкой пищей, имеется хоботок, но построен он по-разному в разных отрядах. У бабочек хоботок образован нижними челюстями, у мух - нижней губой, у пчел - нижними челюстями и нижней губой, а у комаров, клопов - всеми ротовыми частями.
Грудь насекомых состоит из трех сегментов: передне-, средне- и заднегруди. Это локомоторный отдел тела. К груди причленяются три пары ног, а у крылатых насекомых еще и две пары крыльев. Каждый грудной сегмент имеет сложное строение из чередующихся склеритов и участков мембран, что обеспечивает его прочность и подвижность. Различают спинные склериты (тергиты), брюшные (стерниты) и боковые (плевриты). Хитиновые покровы груди образуют внутренние впячивания - эндоскелет, к которым прикрепляются мышцы ног и крыльев.
Конечности насекомых состоят из пяти отделов: базального членика - тазика (соха), вертлуга (trochanter), бедра (femur), голени (tibia) и лапки (tarsus), состоящей из нескольких члеников, последний из которых заканчивается одним или двумя коготками (рис. 319).
В зависимости от выполняемой функции ноги насекомых могут быть бегательными, ходильными, копательными, хватательными, прыгательными, плавательными, присасывательными. Каждый тип конечностей характеризуется спецификой морфологических особенностей.
Ноги насекомых представляют собой самые совершенные органы движения среди конечностей всех членистоногих. Причленение ног у насекомых смещено к брюшной стороне, что определяет приподнятость тела над субстратом. Многочлениковые ноги насекомых с 4-5 суставами обеспечивают успех передвижения по сложному микрорельефу. Большим преимуществом ног насекомых является образование лапки, состоящей из 3-5 члеников. Лапка функционально соответствует стопе сухопутных позвоночных животных. Опушенность нижней стороны лапок, а также особые присоски под коготками у некоторых видов дают насекомым возможность передвигаться не только по вертикальным поверхностям, но и вниз спиной и вверх ногами (например, движение мухи по потолку).
В пределах класса насекомых прослеживается переход от относительно примитивных конечностей, едва приподнимающих тело над субстратом,
434
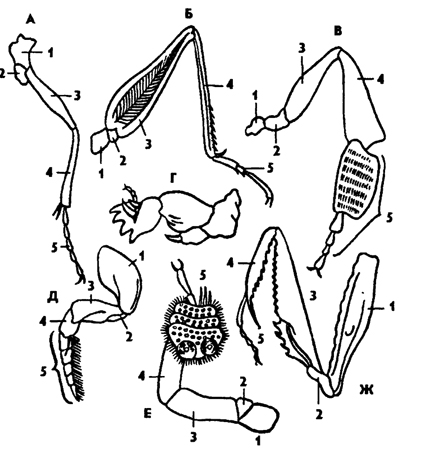
Рис. 319. Конечности насекомых (из Натали): А - бегательная, Б - прыгательная, B - собирательная с корзиночкой (пчела), Г - копательная, Д - плавательная, Е - присасывательная, Ж - хватательная; 1 - тазик, 2 - вертлуг, 3 - бедро, 4 - голень, 5 – лапка
например у первичнобескрылых чешуйниц, до специализированных и совершенных ног у многих жуков, двукрылых.
Исходный тип ног у насекомых - это ходильные и бегательные. От них прослеживаются морфологические переходы к специализированным типам ног - копательным, хватательным, плавательным и др.
Крылья - характерное приспособление к полету у насекомых. Обычно крыльев две пары, которые расположены на средне- и заднегруди. Они представляют собой складки стенки тела. Крыло состоит из двух слоев кожи, покрытых кутикулой, и узкой полости между ними. В толще крыла проходят каналы с трахеями, нервами и лакунами полости - миксоцеля. Они образуют жилки. Крылья закладываются мягкими складками на стадии куколки. У насекомых, выходящих из куколки, по жилкам крыльев нагнетаются гемолимфа и воздух по трахеям. В результате крылья расправляются. После затвердевания покровов крыло омертвевает и представляет собой пластинку, приводимую в движение специальной крыловой мускулатурой. Жилки крыльев выполняют функцию опорной конструкции. Различают продольные жилки: костальную, субкостальную, радиальную, медиальную, кубитальную, анальную и югальную (рис. 320). Продольные жилки, кроме костальной, могут ветвиться. Между ними часто имеются поперечные жилки, которые ограничивают участки крыла, называемые ячейками. Ячейки имеют специальные названия, как и жилки. Строение крыльев и жилкование - важный
435
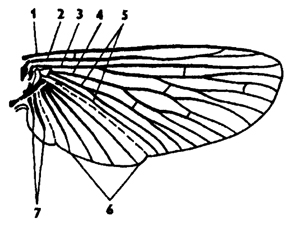
Рис. 320. Схема строения крыла насекомого (из Кенигсмена): 1 - костальная жилка, 2 - субкостальная, 3 - радиальная, 4 - медиальная, 5 - кубитальная, 6 - анальная, 7 - югальные жилки
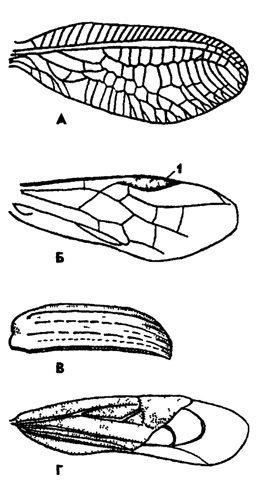
Рис. 321. Типы крыльев у насекомых (по Веберу): А - сетчатое (переднее крыло золотоглазки), Б - перепончатое (переднее крыло пилильщика), B - жесткое надкрылье жука, Г - полужесткое крыло клопа; 1 - птеростигма
систематический признак. По особенностям жилкования крыльев можно определять отряды, семейства, роды и виды насекомых. Разнообразие типов крыльев показано на рис. 321.
У самых примитивных насекомых крыльев нет. Это первичнобескрылые насекомые. К ним относятся чешуйницы. Большинство насекомых имеет две пары крыльев. В некоторых отрядах передняя пара крыльев превращается в жесткие пластинки - надкрылья, прикрывающие в покое задние функционирующие крылья (жуки). В этом случае насекомые летают на задней паре крыльев. Их относят к "заднемоторным".
Мухи и комары, наоборот, летают на передней паре крыльев ("переднемоторные"), а задняя пара крыльев у них редуцирована и превратилась в жужжальца. У высших насекомых из отрядов перепончатокрылых и бабочек передние и задние крылья скрепляются между собой и образуют как бы только два функциональных крыла.
По способу прикрепления крыльев к телу и типу движения среди насекомых различают древнекрылых насекомых и новокрылых. У первых крылья движутся в одной вертикальной плоскости (вверх-вниз). Крылья у второй группы насекомых при полете вращаются вокруг оси и их вершина описывает восьмерку (пчелы). У некоторых насекомых крылья редуцированы, и их называют вторичнобескрылыми. К ним относятся многие паразиты: вши, клопы, блохи. Но бескрылость развивается и по другим экологическим причинам Бескрылы пещерные, некоторые почвенные насекомые, обитатели гнезд муравьев и термитов и т. д.
436
Полет обеспечивается движением крыловой мускулатуры и особенностями причленения крыльев к тергитам. Крыло представляет как бы двуплечий рычаг с разной длиной плеч. Недалеко от основания крыла имеется вырост боковой пластинки тела, на который крыло опирается. Дорзовентральные пучки мышц приводят к опусканию тергита, который давит на короткое плечо и приводит к подъему пластинки крыла. И наоборот, подъем тергита приводит к опусканию крыла. Мышцы, прикрепленные к основанию крыла, способны его поворачивать вдоль продольной оси. При движении крыльев вперед и вниз возникает подъемная сила, а при движении назад и вверх создается пропеллирующий эффект. В первом положении крылья располагаются горизонтально, а при втором - вертикально.
Скорость взмахов крыльев в полете различна: у бабочек 5-9 взмахов в секунду, у пчелы около 200, у комаров-звонцов 1000. Скорость полета также сильно варьирует: у пчелы 3 м/с, у бражников 15 м/с. Длительные перелеты способны делать стрекозы и некоторые бабочки. Например, олеандровый бражник летом развивается на юге - в Крыму, на Кавказе, а осенью может совершать миграции на север до Прибалтики.
Проблема происхождения крыльев обсуждается учеными давно. Наиболее принятая концепция происхождения крыльев из паранотальных выростов кожи на грудных сегментах. У некоторых ископаемых насекомых паранотальные выросты образовывались на трех грудных сегментах и служили для парения при прыжках с растений вниз. Из таких разросшихся выростов в дальнейшем могли образовываться крылья.
Брюшко - третий отдел тела насекомых - основное вместилище внутренних органов. Число сегментов брюшка варьирует. Максимальное число сегментов - 10 характерно для низших отрядов насекомых и для личинок насекомых с полным превращением. В целом наблюдается тенденция к олигомеризации сегментов. У некоторых мух число сегментов равно всего лишь 4-6. Брюшко насекомых лишено конечностей. Однако у некоторых имеются рудименты конечностей, выполняющие другие функции. У чешуйниц (Thysanura) имеются на всех сегментах придатки - грифельки, на которых брюшко волочится по субстрату, как на полозьях санок. Одна пара задних грифельков имеется у тараканов. Членистые придатки заднего сегмента - церки - также производные конечностей. Имеются доказательства, что яйцеклады многих насекомых представляют собой видоизмененные конечности трех последних сегментов, например яйцеклад у кузненечиков, сверчков (рис. 322).
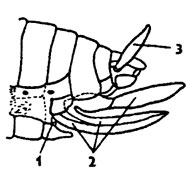
Рис. 322. Схема строения яйцеклада насекомого (из Бей-Биенко): 1 – половое отверстие, 2 - створки яйчиков цеклада, 3- церки
437
Кожно-мускульная система. Кожа насекомых представлена одним слоем клеток - гиподермой. Снаружи гиподерма выделяет кутикулу, а с внутренней стороны она подстилается базальной мембраной соединительнотканной природы (рис. 250).
Кутикула состоит из трех слоев. Наружный слой - эпикутикула - очень тонкий и содержит в своем составе липоиды и воскоподобные вещества. Наиболее хорошо развита эпикутикула у открытообитающих насекомых и слабее у скрытоживущих. Этот слой защищает тело насекомых от высыхания. Второй слой кутикулы - экзокутикула - имеет столбчатое строение, а третий - эндокутикула - слоистой структуры. Эти слои кутикулы во много раз толще эпикутикулы и представляют основу наружного скелета. Экзокутикула и эндокутикула в основном состоят из хитина и пропитаны белками, задубленными фенолами.
Хитиновая кутикула образует серию внутренних выростов - эндоскелет (тенторий), к которым изнутри прикрепляются мышцы и внутренние органы. Производными покровов могут быть: скульптурные образования - (бугорки, ямки, шипы) и более сложные - структурные (щетинки, волоски, шпоры). Волоски у насекомых в некоторых случаях преобразованы в чешуйки, например у бабочек на крыльях.
В гиподерме насекомых много желез. Это могут быть восковые железы, как у пчелы; пахучие, как у клопов; ядовитые, как у некоторых гусениц. Слюнные и паутинные железы также кожного происхождения.
Окраска покровов насекомых зависит от пигментов, которые могут содержаться в кутикуле или гиподерме. Основные пигменты насекомых - меланины, окраска которых варьирует от желтых и бурых до черных. Кроме того, распространены пигменты - каротиноиды и птерины желтых и красных тонов. Переливчатая и металлическая окраска зависит от структуры кутикулы и обусловлена интерференцией света.
Окраска насекомых имеет важное защитное значение. У одних насекомых наблюдается покровительственная окраска под цвет фона, на котором они обитают (зеленый кузнечик - на траве), у других, как у ядовитых и несъедобных видов, - предостерегающая окраска (красная божья коровка с черными пятнами), третьи имитируют окраску (мимикрия) под защищенные виды (муха-пчеловидка похожа на жалящую пчелу).
Мышечная система. Мышечная система насекомых дифференцирована. Различают скелетные мышцы, приводящие в движение тело и его придатки, и внутренностные, входящие в состав внутренних органов. Почти все мышцы насекомых поперечнополосатые.
Относительная сила мышц насекомых велика: они могут передвигать груз, в 14-25 раз превышающий массу их тела. Прыгающие насекомые (саранча, блохи) могут прыгать на расстояние, в сотни и тысячи раз превышающее длину их тела. Частота сокращений мышц может достигать
438
1000 раз в секунду. Интенсивность работы мышц насекомых объясняется высокой скоростью химических процессов в мышцах при условиях трахейного дыхания и способностью мышц отвечать несколькими сокращениями на один нервный импульс.
Пищеварительная система насекомых, как у всех членистоногих, состоит из трех отделов (рис. 323). Передняя кишка эктодермальная и выстлана кутикулой. У большинства видов этот отдел подразделяется на глотку, пищевод, нередко расширяющийся в зоб и мускульный желудок (провентрикулюс). К переднему отделу кишечника примыкают 1-2 пары слюнных желез кожного происхождения. Первая пара желез вырабатывает пищеварительные ферменты. Вторая пара желез может видоизменяться в шелкоотделительные или паутинные железы - у гусениц бабочек. Протоки каждой пары слюнных желез соединяются в непарный канал, который открывается у основания нижней губы под выступом глотки - гипофаринксом. В переднем отделе кишечника происходят механическая обработка пищи и частичное переваривание под действием пищеварительных ферментов слюнных желез. В зависимости от типа питания строение этого отдела кишечника сильно варьирует. Так, у пчел имеется "медовый желудок" - слепой вырост зоба, в котором пчела накапливает мед, чтобы потом отложить его в соты. У некоторых мух зоб представляет слепой резервуар, связанный с пищеводом тонким каналом. Желудок растительноядных насекомых с крупными хитиновыми зубцами внутри, а у некоторых хищников с цедильным аппаратом из длинных волосков.
Средняя кишка энтодермальная; в ней происходит переваривание и всасывание пищи. На границе с передней кишкой средняя кишка часто образует слепые выросты - пилорические придатки, которые функционируют как пищеварительные железы, а также служат для всасывания переваренной пищи. В средней кишке непрерывно выделяется особая пленка - перитрофическая мембрана, обволакивающая пищу. Мембрана защищает стенки кишечника от механических повреждений и облегчает процесс проникновения пищеварительных ферментов в пищевой комок.
У разных насекомых в зависимости от пищевой специализации вырабатываются различные пищеварительные ферменты. Так, у плотоядных видов преобладают протеолитические ферменты, а у растительноядных видов переваривание клетчатки осуществляется за счет кишечных симбионтов (простейших и бактерий), способных выделять фермент целлюлазу. Например, в кишечнике у термитов, питающихся древесиной, обитает несколько десятков видов симбионтов, обеспечивающих переваривание пищи. Если кишечник термитов освободить от симбионтов (дефаунизировать), то они погибают.
На границе среднего и заднего отделов кишки впадают мальпигиевы сосуды (от 2 до 150) эктодермального происхождения.
439
Задняя кишка иногда дифференцируется на несколько участков: тонкую, толстую и прямую. В стенках прямой кишки находятся продольные валики - ректальные железы, которые обеспечивают наиболее полное всасывание влаги из экскрементов и экскретов, выделяемых через анальное отверстие.
Органы выделения насекомых, как у всех трахейных, представлены мальпигиевыми сосудами, впадающими в заднюю кишку. Основной продукт выделения - кристаллы мочевой кислоты. У насекомых, живущих в воде или во влажных местах, мальпигиевых сосудов много (до 150), так как они выполняют дополнительную функцию осморегуляции. У видов, обитающих в аридных условиях, мальпигиевых сосудов всего 1-2 пары, а их слепые концы нередко прирастают к прямой кишке, что усиливает обратное всасывание воды из мальпигиевых сосудов в гемолимфу и более надежно обеспечивает экономию влаги в организме.
Дополнительно выделительную функцию выполняют почки накопления: клетки жирового тела и перикардиальные клетки, скопления которых располагаются вокруг сердца. Частично продукты обмена откладываются в покровную кутикулу, а затем при линьке личинок удаляются вместе с экзувием - сброшенной кутикулой.
Жировое тело у насекомых сильно развито и выполняет несколько функций. Главная функция жирового тела - это накопление запаса питательных веществ (жиров, белков и гликогена) и метаболической влаги. За счет этих энергетических веществ происходит интенсивный рост насекомых и сложный метаморфоз. Благодаря запасам питательных веществ насекомые могут долго обходиться без пищи и воды. Особенно это важно при переживании неблагоприятных условий сезона: зимовки или летней засухи. Некоторые паразиты, например постельные клопы, могут голодать несколько месяцев и жить за счет жирового тела. Дополнительной функцией жирового тела является накопление продуктов обмена (почка накопления). В клетках жирового тела накапливаются соли мочевой кислоты и другие экскреты. У некоторых насекомых, например жуков-светляков, часть жирового тела видоизменяется в орган свечения. В нем содержится вещество люцеферин, способное к свечению.
Кровеносная система насекомых незамкнутая. Кровь - гемолимфа - свободно циркулирует по полости тела, омывая все органы. Ее приводит в движение главный пульсирующий орган - сердце. Сердце насекомых расположено на спинной стороне и имеет форму мускулистой трубки, разделенной на камеры. На заднем конце сердце слепо замкнуто. Камер может быть восемь, но их число может сокращаться до одной. Каждая камера имеет пару боковых отверстий - остий с клапанами, обеспечивающими односторонний ток крови из полости тела в сердце. Между камерами имеются клапаны, открывающиеся только вперед (рис. 324).
440
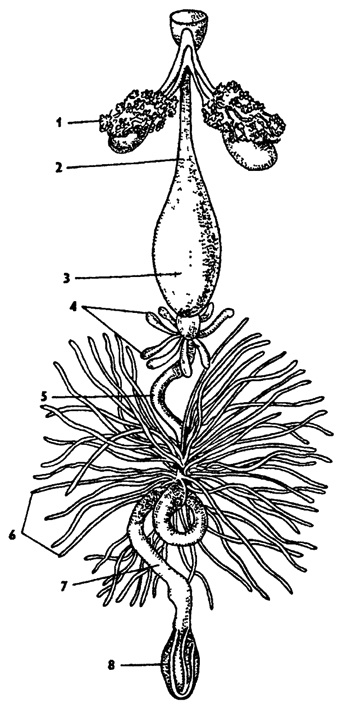
Рис. 323. Пищеварительная система черного таракана (по Веберу): 1 - слюнные железы, 2 - пищевод, 3 - зоб, 4 - пилорические придатки, 5 - средняя кишка, 6 - мальпигиевы сосуды, 7 - задняя кишка, 8 - прямая кишка
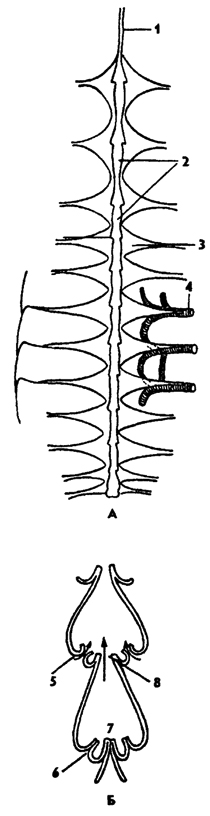
Рис. 324. Сердце таракана (А) и строение камер сердца (Б) (из Натали): 1 - аорта, 2 - камеры сердца, 3 - крыловидные мышцы сердца, 4 - трахея, 5 - остии камеры сердца открыты, 6 - остии камеры сердца закрыты, 7 - клапан между камерами закрыт, 8 - клапан между камерами открыт
441
Поэтому волна сокращений сердца от задней камеры к передней обеспечивает движение крови вперед. Сердце подвешено на коротких мышечных тяжах к спинной стенке тела (рис. 325). Снизу к важдой камере сердца прикрепляется пара крыловидных мышц, похожих на крылья или веера (рис. 324, А). Эти мышцы соединяют сердце с боковыми частями тергитов. Горизонтальная перегородка, образованная крыловидными мышцами и соединяющей их пористой мембраной, называется перикардиальной диафрагмой (рис. 325). Она отделяет околосердечную - перикардиальную - полость, или синус, от остальной полости тела. При расслаблении крыловидных мышц диафрагма несколько опускается, и кровь устремляется в околосердечную полость, а при их сокращении кровь поступает через остии в сердце. Последовательное сокращение камер сердца гонит кровь вперед в головную аорту (рис. 324, Б). Кровь из аорты изливается в полость тела. Имеются дополнительные пульсирующие органы в груди, конечностях, обеспечивающие ток крови в придатках. На брюшной стороне тела над нервной цепочкой у некоторых насекомых образуется еще одна - периневральная диафрагма с мышцами. Ее сокращение обеспечивает движение крови назад по брюшной полости и по периневральному синусу. Таким образом, пульсация сердца, сокращение диафрагм и некоторых дополнительных пульсаторов способствуют циркуляции крови в теле насекомых.
Кровь насекомых состоит из плазмы и кровяных телец - гемоцитов, среди которых различают фагоциты, лейкоциты и амебоциты.
Функции крови состоят в снабжении органов питательными веществами и переносе продуктов обмена к органам выделения. Кроме того, кровь насекомых выполняет гуморальную регуляцию жизненных процессов. В связи с сильно развитой трахейной системой кровь насекомых практически не выполняет дыхательную функцию. Однако у некоторых водных личинок насекомых - комаров (Chironomus), которых обычно называют "мотылем", в крови имеется красный дыхательный пигмент, близкий по формуле к гемоглобину. Это свидетельствует о том, что в условиях дефицита кислорода кровь может дополнительно выполнять и дыхательную фукнцию, обеспечивая транспорт газов. Кроме этого, кровь насекомых определяет тургор тела, что особенно важно для личинок с мягкими покровами. Под давлением гемолимфы расправляются крылья у молодого насекомого и разрываются покровы при линьке.
У некоторых насекомых кровь может быть ядовитой, что имеет защитное значение. Например, у божьей коровки и жуков-нарывников кровь обладает едкими свойствами. Эти насекомые в случае опасности способны выбрызгивать кровь из мест сочленения сегментов и ног. Это произвольное кровопускание - аутогеморрагия. Яркая окраска таких насекомых предупреждает хищников о том, что они ядовиты и несъедобны.
442
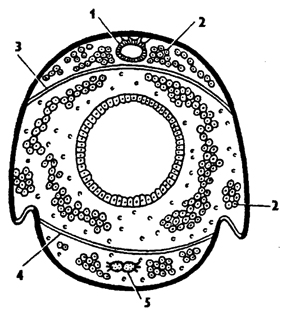
Рис. 325. Поперечный разрез через тело насекомого (из Бей-Биенко): 1 - сердце, 2 - жировое тело, 3 - перикардиальная диафрагма, 4 - периневральная диафрагма, 5 - брюшная нервная цепочка
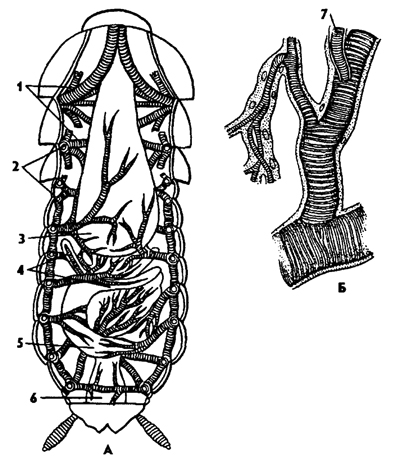
Рис. 326. Трахейная система черного таракана Blatta orientalis (А) и участок трахеи (Б) (из Догеля): 1 - трахейные стволы, 2 - стигмы, 3 - зоб, 4 - пилорические придатки, 5 - средняя кишка, 6 - задняя кишка, 7 - тенидии
Дыхательная система насекомых представлена трахейной системой (рис. 326, А). Трахеи представляют собой глубокие впячивания покровов, открывающиеся наружу дыхальцами - стигмами. Изнутри трахеи выстланы тонкой кутикулой - интимой, которая образует спиральные утолщения - тенидии (рис. 326, Б). Тенидии препятствуют спаданию трахей. Трахеи ветвятся, опутывают все внутренние органы. Концевые ветви трахей заканчиваются звездчатой трахейной клеткой, от которой отходят тончайшие трахейные трубочки - трахеолы, проникающие даже внутрь клеток окружающих тканей.
Трахейная система насекомых сильно развита. Максимально бывает 10 пар дыхалец, из которых две пары расположены на средне- и заднегруди, а остальные восемь пар на последующих брюшных сегментах. От дыхалец отходят короткие поперечные трахеи, впадающие в боковые трахейные стволы, от которых отходят многочисленные ветвящиеся трахеи. Иногда трахеи образуют расширения - воздушные мешки, которые уменьшают удельный вес тела и улучшают вентиляцию в трахейной системе. Например, у майского жука множество воздушных мешков, что облегчает его удельный вес. У хорошо летающих насекомых - комнатной мухи и пчелы - воздушные мешки занимают большую часть полости тела.
Трахейная система с дыхальцами называется открытой. Имеется большое разнообразие открытых трахейных систем - от типичной с
443
10 парами дыхалец до системы с одной парой дыхалец на конце брюшка, как, например, у водных личинок мух-пчеловидок - "крысок" с длинной дыхательной трубкой, выступающей из воды.
У ряда водных личинок насекомых трахейная система может быть закрытой, без дыхалец. В этом случае у насекомых развиваются трахейные жабры - выросты кожи с густой сетью трахей внутри. Это дает возможность водным насекомым дышать кислородом, растворенным в воде. Газообмен происходит через кожу. Растворенный в воде кислород переходит в трахейную систему в газообразном состоянии. Трахейные жабры у личинок поденок представляют собой парные листовидные выросты на брюшных сегментах (рис. 327), а у личинок стрекоз-стрелок три хвостовые пластинки. У личинок стрекозы-коромысло имеются внутренние трахейные жабры в ректальном отделе кишки.
В отличие от других членистоногих у насекомых транспорт кислорода к тканям выполняется в основном только при помощи трахей, без участия кровеносной системы.
Нервная система насекомых, как у всех членистоногих, построена по типу брюшной нервной цепочки. У высших отрядов насекомых нервная система может достигать высокого уровня развития и дифференциации.
Головной мозг, или парный надглоточный узел, состоит из трех отделов: прото-, дейто- и тритоцеребрума (рис. 328). Протоцеребрум иннервирует акрон и размещающиеся на нем глаза, дейтоцеребрум - усики, тритоцеребрум - верхнюю губу. На протоцеребруме развиты грибовидные тела - важные ассоциативные центры мозга, к которым подходят нервы от органов зрения. Чем сложнее поведение насекомых, тем сильнее развит у них головной мозг и грибовидные тела, например у рабочих особей пчел и муравьев.
Головной мозг связан окологлоточными коннективами с подглоточным ганглием, который состоит из слившихся трех пар ганглиев, иннервирующих ротовой аппарат и передний отдел кишечника.
Брюшная нервная цепочка иннервирует грудь и брюшко. Максимально в ее состав входит 11-13 пар ганглиев: три грудных и 8-10 брюшных. В пределах класса наблюдается тенденция к олигомеризации ганглиев брюшной нервной цепочки. Так, например, у тараканов брюшная нервная цепочка представлена восемью ганглиями; у жуков-вертячек ганглии брюшного отдела сливаются в общий узел, а у многих мух грудные и брюшные ганглии цепочки сливаются в один грудной узел.
С центральной нервной системой связана система симпатических нервов, иннервирующих внутренние органы.
Эндокринные железы насекомых представлены нейросекреторными клетками мозга, кардиальными, прилежащими телами и переднегрудными железами (рис. 329).
444
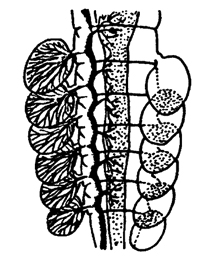
Рис. 327. Трахейные жабры личинки поденки (из Шванвича)
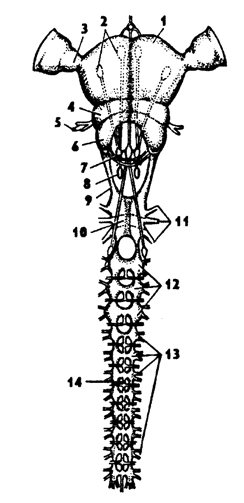
Рис. 329. Нейро-гуморальная система у насекомых (из Догеля): 1 - головной мозг, 2 - нейро-секреторные клетки, 3 - кардиальное тело, 4 - прилежащее тело, 5 - проторакальная железа, в - гонада, 7 - линька кутикулы; АГ - активационный гормон, ЮГ - ювениальныи гормон, ЛГ - линочный гормон
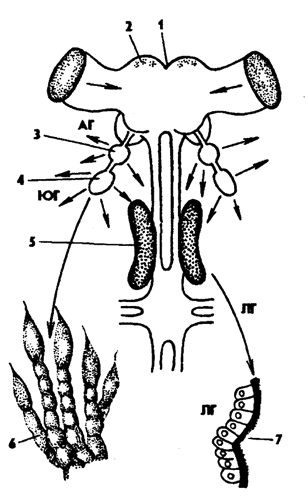
Рис. 328. Нервная система насекомых (из Вюрбаха): 1 - протоцеребрум, 2 - нейросекреторные клетки, 3 - грибовидное тело, 4 - дейтоцеребрум, 5 - антеннальный нерв, 6 - тритоцеребрум, 7 - кардиальные тела, 8 - прилежащие тела, 9 - окологлоточная коннектива, 10 - подглоточный ганглий, 11 - нервы конечностей, 12 - фудные ганглии, 13 - брюшные ганглии, 14 - непарный нерв симпатической системы
445
В состав центральной нервной системы входят нейросекреторные клетки, из которых нейросекреты по аксонам поступают в кардиальные, прилежащие тела, переднегрудные железы, а из них выделяемые гормоны поступают в гемолимфу. Нейросекреты регулируют деятельность эндокринных желез, выделяющих гормоны, которые регулируют рост, линьки, развитие организма, обмен веществ.
Прилежащие тела располагаются над передней кишкой позади головного мозга. Кардиальные тела близко примыкают к прилежащим телам и расположены впереди последних. Они связаны нервами с головным мозгом. Прилежающие тела секретируют у личинок ювенильный гормон, который способствует личиночному развитию и препятствует метаморфозу - превращению в куколку. В имагинальном состоянии насекомых ювенильный гормон стимулирует созревание половых продуктов. Кардиальные тела регулируют обменные процессы в организме. Переднегрудные железы более крупные, располагаются по бокам переднегрудных ганглиев, с которыми связаны нервами. Эти железы присутствуют только на преимагинальных стадиях развития, а на стадии имаго эти железы дегенерируют. Секрет переднегрудных желез линочный гормон (экдизон) стимулирует линьку.
Деятельность желез внутренней секреции взаимосвязана с нервной системой. Таким образом, у насекомых имеется нейрогуморальная регуляция жизненных процессов. Эта система регуляции зависит также и от факторов внешней среды, например от сезонных явлений в природе. Так, например, короткий световой день подавляет нейросекрецию, а за ней и деятельность прилежащих тел и переднегрудных желез. Это приводит к остановке развития и переходу к состоянию диапаузы перед зимовкой. И наоборот, при длинном световом дне активизируется действие экдизона и ювенильного гормона, что приводит к активному росту личинок.
Органы чувств насекомых достигают высокой степени совершенства. Возможности их чувствующего, или сенсорного, аппарата нередко превосходят таковые у высших позвоночных животных и человека. Например, пчелы различают нам недоступный ультрафиолетовый цвет; многие мухи различают поляризованный свет. Органы слуха насекомых способны воспринимать не только звуковые колебания, но и любые колебания среды. Так, многие ночные бабочки реагируют на ультразвуки. Доказано сейсмическое и магнетическое чувство у насекомых. Химические анализаторы насекомых также обладают высокой чувствительностью. Методом мечения было выявлено, что самцы некоторых бабочек улавливают аттрактанты самок с привлекающим запахом на расстоянии 11 км. Комнатная муха различает присутствие сахара в воде в 2000 раз меньших концентрациях, чем человек. Тончайшие восприятия влажности воздуха наблюдаются у многих приводных видов. При полете они чувствуют местонахождение водоемов на большом расстоянии.
446
У насекомых органы чувств подразделяются на механорецепторы, хеморецепторы, термо- и гигрорецепторы, фоторецепторы.
Элементарным элементом всех органов чувств насекомых являются сенсиллы. Сенсилла состоит из одной или нескольких чувствующих клеток, каждая из которых имеет чувствующий периферический отросток и центральный отросток, ведущий к нервной системе. На конце чувствующего отростка клеток имеется видоизмененный жгутик без центральных фибрилл, заключенный в кутикулярную трубочку (рис 330). Сенсиллы насекомых разнообразны по строению в зависимости от выполняемой функции.
Механорецепторы воспринимают механические раздражения. Они могут обладать контактным или дистанционным восприятием. Это органы осязания, слуха и сейсмического чувства.
Органы осязания представлены сенсиллами с контактным восприятием. Особенно они многочисленны на усиках насекомых. Членики усиков густо покрыты тонкими подвижными волосками, у основания которых расположены осязательные сенсиллы. Простейшая осязательная сенсилла состоит из одной клетки с центральным и периферическим отростками (рис 330). Конец жгутика периферического отростка подходит к основанию чувствующего волоска и все осязательные раздражения передает чувствующей клетке.
Органы слуха и сейсмического чувства обладают дистантным восприятием различного рода колебаний среды. Сенсиллы этих органов чувств специализированы и называются сколпофорами. У них периферический отросток чувствующей клетки со жгутиком, заключенным в кутикулярную трубочку, окружен обкладочными клетками с чехликом из фибрилл (рис 331). Этот фибриллярный чехол называется сколопоидным тельцем. Чувствующий жгутик сверху прикрыт шапочковидной клеткой, за счет которой образуется куликулярная трубочка вокруг дистального конца жгутика.
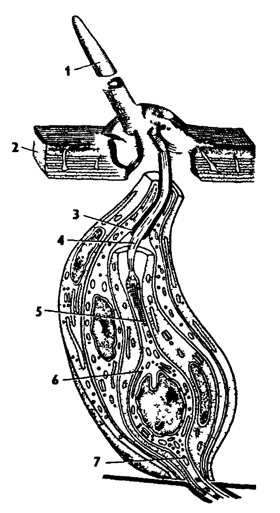
Рис. 330. Строение механорецепторной сенсиллы насекомых (по В Иванову): 1 - чувствительный волосок, 2 - кутикула, 3 - кутикулярная трубочка, 4 - видоизмененный жгутик, 5 - периферический отросток чувствительной клетки, 6 - чувствительная клетка, 7 - центральный отросток чувствительной клетки
447
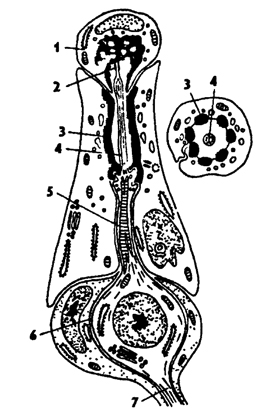
Рис. 331. Строение хордотональной сенсиллы насекомых (по В Иванову): 1 - шапочковая клетка, 2 - кутикулярная трубочка, 3 - сколопоидное тельце, 4 - видоизмененный жгутик, 5 - периферический отросток чувствительной клетки, 6 - чувствительная клетка, 7 - центральный отросток чувствительной клетки
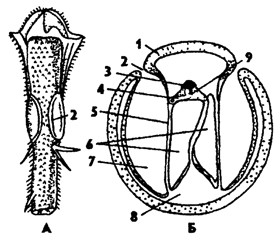
Рис. 332. Тимпанальный орган (слуха) кузнечика: А - передняя голень, Б - поперечный разрез через голень в области тимпанального органа (по Швабе); 1 - кутикула, 2 - слуховая щель, 3 - шапочковая клетка, 4 - сколпофор, 5 - барабанная перепонка, 6 - трахеи, 7 - барабанная полость, 8 - полость ноги, 9 - сколопоидное тельце
Сколпофоры входят в состав хордотональных органов, или органов слуха. Часть этих органов примыкают к внутренней поверхности кутикулы или находятся на особых мембранах, натянутых между двумя участками кутикулы. Хордотональные органы у разных насекомых располагаются на разных частях тела: брюшке, усиках, ногах, крыльях. Эти органы воспринимают механическое напряжение, внутреннее давление, механические вибрации. Они способствуют регуляции положения тела в пространстве, особенно во время полета.
Сколпофоры входят в состав тимпанальных органов, или органов слуха. В схеме тимпанальный орган состоит из кутикулярной мембраны, с которой связаны скопления сколпофоров. Органы слуха особенно хорошо развиты у насекомых, у которых имеются органы звука.
У кузнечиков органы слуха находятся на передних голенях (рис. 332). Парные боковые щели ведут в карманообразные углубления с тонкими внутренними стенками - мембранами. Мембраны с двух сторон примыкают к продольным стволам трахей. Вибрация мембран резонирует в трахеях. К мембранам и трахеям примыкают группы сколпофоров, от которых отходит слуховой нерв.
У саранчовых органы слуха находятся на первом сегменте брюшка, у певчих цикад - на груди у основания брюшка. Органы слуха имеются лишь у тех насекомых, которым свойственна звуковая сигнализация.
Функцию органов звука у самцов кузнечиков и сверчков выполняют передние крылья. На левом крыле имеется выступающая зубчатая жилка - "смычок", а на правом находится резонатор - "зеркальце",
448
представляющее округлую площадку крыла, ограниченную выступающей жилкой. У самцов саранчовых роль смычка выполняют зубчики на внутренней стороне задних бедер. Звук у них издается при трении бедер о передние надкрылья с выступающей радиальной жилкой. У певчих цикад тимпанальные органы (слуха) играют роль и органов звука. Мембрана тимпанальных органов может производить вибрацию, вызываемую сокращением особых мышц.
Хеморецепторы воспринимают химические раздражения. Может быть контактное восприятие или дистантное. К хеморецепторам относятся органы вкуса и обоняния.
Обоняние - это дистантная хеморецепция. Органы обоняния могут располагаться на разных частях тела насекомых. Особенно много обонятельных сенсилл на усиках. Они могут иметь вид выступающих тонкостенных пузырьков, конусов, щелей, ямочек, пластинок. Так, у рабочей пчелы на каждом усике до б тыс. пластинчатых сенсилл. У мух они собраны в особую ямку на третьем членике усиков.
Обонятельные сенсиллы по своему строению несколько отличаются от других. Каждая обонятельная сенсилла состоит из группы чувствующих клеток, периферические отростки которых образуют пучок жгутиков, заключенный в широкую кутикулярную трубочку. Окончания жгутиков подходят к кутикулярному образованию сенсиллы - пузырьку, пластинке или ямке, которые пронизаны тончайшими порами (рис. 333). Таким образом, химические раздражители через поры действуют на жгутики чувствующих клеток, от которых отходят центральные отростки к нервной системе.
Обоняние имеет большое значение в жизни насекомых. По запаху они отыскивают пищу, места для откладки яиц, а также особей другого пола. Многие насекомые выделяют привлекающие вещества - аттрактанты. Чувствительность обонятельных сенсилл особенно высока на распознавание аттрактантов. Самцы бабочек распознают самок с расстояния 3-9 км.
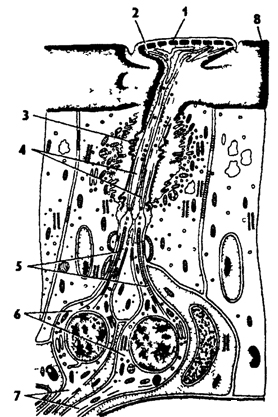
Рис. 333. Обонятельная сенсилла насекомых (по В Иванову): 1 - кутикулярная пластинка, 2 - поры, 3 - кутикулярная трубочка, 4 - видоизмененный жгутик, 5 - периферический отросток чувствительной клетки, 6 - чувствительные клетки, 7 - центральный отросток чувствительной клетки, 8 – кутикула
449
Органы вкуса - это контактные хеморецепторы. Они представлены вкусовыми сенсиллами с кутикулярными придатками, пронизанными порами. В отличие от обонятельных, сенсиллы вкуса, как правило, включают большее число чувствующих клеток с дифференциацией восприятия вкуса. Одни клетки ощущают горький вкус, другие - сладкий, третьи - кислый. Вкус насекомых не идентичен вкусу у человека. Так, например, пчела не обнаруживает горького вкуса хинина, добавленного в сахарный сироп.
Органы вкуса у мух и бабочек расположены на лапках передних ног, а у жуков на челюстных и губных щупиках.
Термо - и гигрорецепторы способны воспринимать режим влажности и температуры. Действие этих рецепторов может быть контактным и дистантным. Сенсиллы с такой специализацией расположены преимущественно на усиках и щупиках. Они имеют вид кутикулярных ямочек, бугорков. Насекомые, чувствительные к микроклимату, обладают хорошо развитыми гигро- и терморецепторами. Так, например, личинки жуков-щелкунов, обитающие в почве, способны различать относительную влажность воздуха от 99,5% до 100%, т.е. воспринимают разницу в 0,5%.
Фоторецепторы - органы зрения. Зрение вместе с химическим чувством играет ведущую роль в жизни насекомых. Органы зрения насекомых представлены простыми и сложными - фасеточными глазами.
Сложные глаза расположены по бокам головы и состоят из глазков - омматидиев (рис. 334). Их число может достигать нескольких сотен и даже тысяч. Каждый омматидий состоит из оптического и чувствующего отделов (рис. 335). Оптический отдел омматидия состоит из хитиновой прозрачной фасетки в форме шестигранника и расположенного под ней хрустального конуса, который образован четырьмя удлиненными прозрачными клетками. Хрустальный конус по периферии окружен пигментными клеткам. Чувствующая часть омматидия состоит из 4-12 зрительных ретинальных клеток, расположенных радиально. В центре прикосновения ретинальных клеток образуется зрительный стержень - рабдом. Центральные отростки зрительных клеток переходят в зрительный нерв. Ретинальные клетки также окружены пигментными клетками. Таким образом, каждый омматидий заключен в двухъярусный пигментный бокал, изолирующий его от соседних омматидиев.
У насекомых с дневным зрением пигментные клетки омматидиев переполнены пигментом и надежно изолируют соседние глазки. У насекомых с сумеречным зрением в пигментных клетках пигмента меньше, и его зерна способны перемещаться и концентрироваться в верхней части, что способствует проникновению лучей света в соседние омматидий (рис. 335). Насекомые с сумеречным зрением могут видеть и в дневное
450
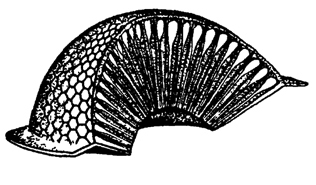
Рис. 334. Сложный глаз насекомого. На разрезе видны омматидии (по Гессе)
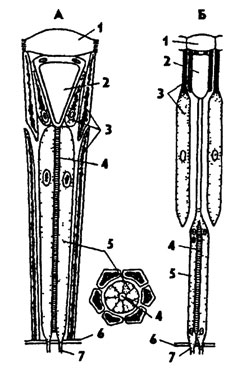
Рис. 335. Строение омматидиев дневных (А) и сумеречных (Б) насекомых (из Мазохина-Поршнякова): 1 - хрусталик, 2 - хрустальный конус, 3 - пигментные клетки, 4 - рабдом, 5 - чувствительные клетки, 6 - базальная мембрана, 7 - центральный отросток чувствительных клеток
время; при этом пигмент в клетках омматидия равномерно распределен и образует оптическую изоляцию омматидиев.
Дневное зрение насекомых называется аппозиционным, а сумеречное - суперпозиционным. При сумеречном зрении наблюдается нарушение эффекта мозаичности зрения.
Для многих насекомых - бабочек, пчел, мух, муравьев известно цветное зрение. Их восприятие цвета отличается от такового у человека. Так, пчелы различают четыре цвета. Первый цвет, который воспринимают пчелы, соответствует красному, желтому и зеленому у человека. Второй цвет - сине-зеленый, третий - сине-фиолетовый и четвертый - ультрафиолетовый.
Простые глазки имеются не у всех насекомых. Они обычно расположены на темени в числе трех, например у пчел, муравьев. По строению они отличаются от омматидиев. Под одной крупной линзой глаза расположено множество групп ретинальных клеток с рабдомами (рис. 335). Кроме того, простые глазки лишены хрустального конуса и иннервируются от переднего отдела протоцеребрума, а не от грибовидных тел.
У личинок насекомых по бокам головы расположены простые глазки - стеммы Их может быть несколько с каждой стороны, от одного до шести. Простые глазки дополняют основную зрительную функцию сложных глаз. Насекомые, у которых имеются только простые глазки, способны лишь различать степень освещенности, что определяет время их суточной активности и избирательность к местам обитания с разной освещенностью.
Половая система. Размножение. Насекомые раздельнополые. У многих видов хорошо выражен половой диморфизм. Так, самцы и самки
451
многих бабочек отличаются по окраске, самцы жуков-оленей и носорогов вооружены выростами на голове и на переднеспинке, самки некоторых видов бескрылы, а самцы крылатые, например у жуков-светляков.
Половая система самцов состоит из парных семенников и семяпроводов, которые впадают в семяизвергательный канал (рис. 336, Б). Кроме этого, к мужской половой системе относятся придаточные железы, секреты которых разбавляют сперму или образуют оболочку сперматофоров. Заканчивается семяизвергательный канал совокупительным органом - эдеагусом, в состав которого входят кутикулярные элементы - гениталии. Строение гениталиев сложное и имеет таксономическое значение. Признаки строения гениталий самца широко используются в современной систематике насекомых.
Половая система самок состоит из двух яичников, яйцевода и непарного влагалища (рис. 336, А). Яичники состоят из яйцевых трубочек, которых может быть 1-100 пар. Во влагалище впадают придаточные железы. У многих видов имеется семяприемник, проток которого открывается во влагалище. У полового отверстия часто имеется яйцеклад. Яйцеклад может быть крупным - саблевидным, как у кузнечиков, сверчков, или коротким, как у саранчи. Хитиновые гениталии самок - важный диагностический признак в систематике многих групп насекомых.
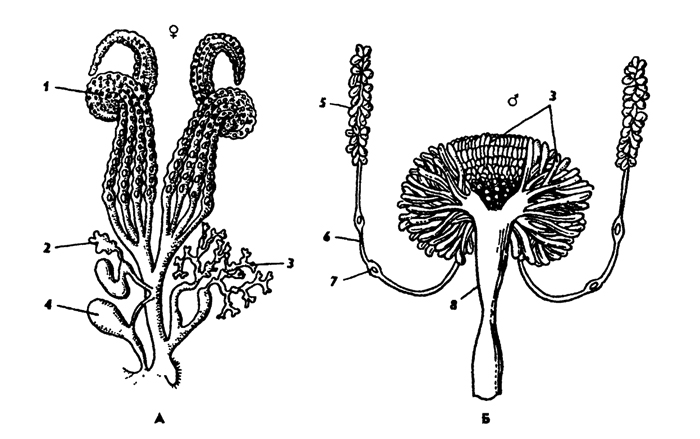
Рис. 336. Половая система самки (А) и самца (Б): 1 - яичник, 2 - семяприемник, 3 - придаточные железы, 4 - совокупительная сумка, 5 - семенник, 6 - семяпровод, 7 - его раздвоение, 8 - семяизвергательный канал (из Догеля)
452
3W.SU ©® 2015