Зоология беcпозвоночных
Класс Ресничные черви, или Планарии (Turbellaria)
Ресничные черви - это преимущественно свободноживущие плоские черви, реже паразитические. Их тело покрыто ресничным эпителием. У большинства видов имеются разнообразные органы чувств. В отличие от паразитических классов плоских червей, у турбеллярий отсутствуют специализированные органы прикрепления, нет сложного жизненного цикла.
Всего известно более 3500 видов ресничных червей. Большинство видов ресничных встречается в морях и пресных водах, меньшее число - во влажных местах на поверхности суши, в почве, и совсем немногие
163
виды являются эктопаразитическими. Представители ресничных червей представлены на рисунке 113.
Тело планарий уплощенное, овальное или удлиненное. На переднем конце тела обычно располагаются органы чувств: глазки, статоцист, обонятельные ямки. Рот - на брюшной стороне тела. Размеры сильно варьируют - от микроскопических до 30- 40 см. Мелкие планарий плавают при помощи ресничного эпителия, а крупные - в основном за счет волнообразных сокращений тела. Ползают планарий по субстрату путем сокращения мускулатуры тела или передвигаются "шагающим" движением, поочередно прикрепляясь то передним, то задним концом тела.
Кожно-мускульный мешок ресничных червей характеризуется наличием ресничного эпителия и нескольких слоев мышц: кольцевых, продольных, диагональных (рис. 114). У некоторых видов эпителий погруженного типа. В эпителии много различных чувствующих и железистых клеток. Обильное выделение слизи помогает скольжению планарий по субстрату. Специфической особенностью эпителиальных клеток планарий является наличие особых палочковидных структур - рабдитов. Рабдиты способны "выстреливать" из клеток наружу, разбухать, превращаясь в ядовитую слизь. Это имеет защитное значение для планарий, а также служит средством поражения добычи.
Пищеварительная система планарий очень разнообразна в пределах класса и варьирует от примитивной - без оформленного кишечника до самой сложной - с ветвистым кишечником. Основные отряды планарий отличаются прежде всего формой кишечника. Рассмотрим типы кишечника некоторых основных отрядов планарий (рис. 115).
У отряда бескишечных планарий (Acoela) имеется рот на брюшной стороне примерно посередине тела. Проглоченная пища попадает в энтодермальную паренхиму, прилегающую к периферическому слою мезодермальной паренхимы (рис. 115, А). Вокруг проглоченного комочка пищи образуется временная пищевая полость, окруженная пищеварительными клетками. В основном они питаются диатомовыми водорослями.
Оформленный и довольно сложный кишечник у многоветвистых планарий (отряд Polycladida). Ротовое отверстие у них, как правило, ближе к заднему концу тела и ведет в складчатую глотку, от которой радиально расходятся множество слепых ветвей энтодермального кишечника (рис. 115, Г).
В отряде трехветвистых планарий (Tricladida) от глотки, расположенной посередине тела, отходят три ветви кишечника. Одна ветвь направлена к переднему концу тела, а две - назад. У отряда прямокишечных (Rhabdocoela) глотка расположена на переднем конце тела и кишечник прямой неветвистый (рис. 115, В).
164
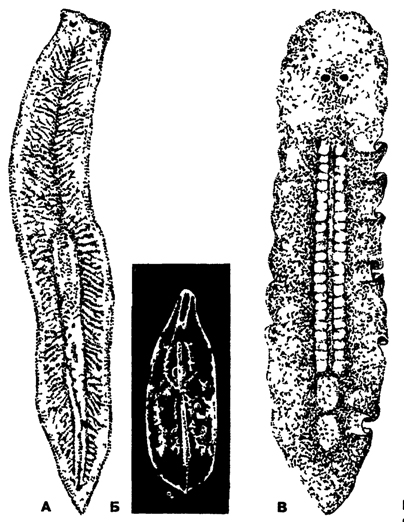
Рис. 113. Планарии (по Стрелкову): А - молочная планария Denflrocoehum lacteum, Б - прямокишечная планария Mesostoma ehrenbergi, В - многоветвистая планария Leptoplana alcinoi
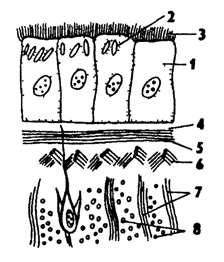
Рис. 114. Схема строения кожно-мускульного мешка: 1 - клетки эпителия, 2 - рабдиты, 3 - реснички, 4 - базальная мембрана, 5 - кольцевые мышцы, 6 - диагональные мышцы 7 - дорзовентральные мышцы, 8 - продольные мышцы
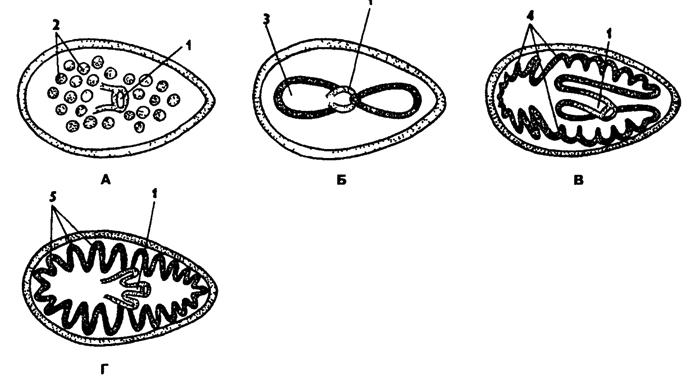
Рис. 115. Типы кишечника у турбеллярий (по Барнсу): А - бескишечные Acoela, Б - большинство отрядов и прямокишечные Rhabdocoela, В - трехветвистые Tricladlda, Г - многоветвистые Polycladida; 1 - глотка, 2 - пищеварительные клетки, 3 - средняя кишка, 4, 5 - ветви средней кишки
165
У планарии с оформленным кишечником большую роль в переваривании пищи выполняют глоточные железы. У многих планарии наблюдается внекишечное пищеварение. Нередко планарии нападают на довольно крупные жертвы (моллюсков, рачков - водяных осликов). В тело жертвы они выделяют пищеварительные ферменты, а затем всасывают полупереваренную пищу.
Нервная система. У бескишечных планарии имеется парный мозговой ганглий на переднем конце тела, от которого отходит сеть нервных тяжей. Это самое примитивное состояние нервной системы у ресничных червей, близкое к диффузной. В других отрядах наблюдается упорядочивание нервной сети и формирование продольных нервных тяжей - коннективов и поперечных - комиссур, при этом образуется решетчатая нервная система - ортогон (рис. 116).
Органы чувств. Кожа планарии усеяна чувствующими клетками с неподвижными длинными ресничками, выполняющими функцию органов осязания и химического чувства. У некоторых видов имеется статоцист, расположенный над мозговым ганглием. Почти у всех планарии присутствуют глаза. Обычно бывает одна пара глаз над мозговым ганглием, но у некоторых может быть несколько десятков глаз, окаймляющих весь передний конец тела (рис. 117). Глаза планарии инвертированного типа (обращенные). Пигментный бокал глаз частично обращен вогнутой частью к поверхности головы. В него погружены длинные изогнутые зрительные клетки, на расширенных концах которых находятся светочувствительные структуры (рис. 118). Свет вначале проходит через тело зрительных клеток и лишь потом попадает в их светочувствительную часть. У других животных с неинвертированными глазами светочувствительные концы зрительных клеток обращены к свету.
Выделительная система планарии протонефридиального типа. Обычно имеется один или два основных канала, от которых отходит множество ветвящихся мелких канальцев, заканчивающихся мерцательными клетками - циртоцитами (рис. 119). На заднем конце тела выделительные каналы открываются наружу выделительными порами. Наиболее сильно развиты протонефридии у пресноводных турбеллярий, что связано с их дополнительной функцией осморегуляции. У бескишечных планарии экскреторную функцию выполняют амебоидные клетки, накапливающие экскреты. Переполненные экскретами клетки удаляются через кожу.
Половая система. Планарии - гермафродиты. В каждой особи имеются женские и мужские половые железы - гонады. У многих видов имеется сложная система половых протоков и дополнительные железы
166
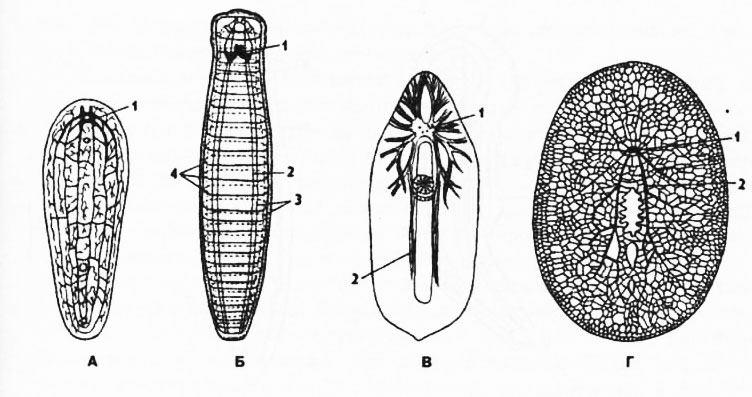
Рис. 116. Нервная система турбеллярий (по Беклемишеву) из разных отрядов: А - Acoela, Б - отряд, близкий к Tndadida, В - Rhabdocoela, Г - Polycladlda; 1 - мозговой ганглий, 2 - брюшные продольные стволы, 3 - боковые и спинные продольные нервные стволы, 4 - комиссуры

Рис. 117. Планария-многоглазка Polycoehs cornuta
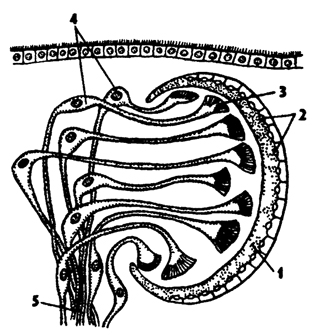
Рис. 118. Схема инвертированного глаза планарий (по Гессе): 1 - пигментный бокал, 2 - ядра клеток пигментного бокала, 3 - светочувствительная часть рецепторных клеток, 4 - ядра рецепторных клеток, 5 - нервные волокна, образующие зрительный нерв
167
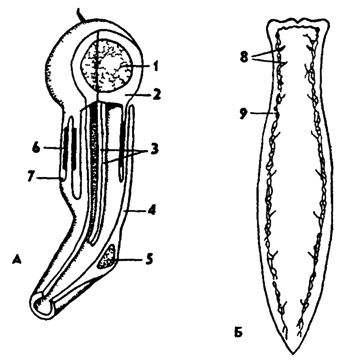
Рис. 119. Протонефридии турбеллярий (по Барнсу): А - клетка с ресничным пламенем, Б - система протонефридиев; 1 - ядро, 2 - терминальная клетка, 3 - жгутики, 4 - трубчатая клетка, 5 - ядро трубчатой клетки, 6 - соединение двух клеток, 7 - отверстия, 8 - нефридиальные поры, 9 - нефридиапьные каналы
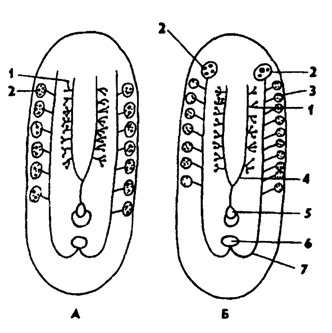
Рис. 120. Схема половой системы турбеллярий (по Барнсу): А - архоофоры, Б - неофоры; 1 - семенники, 2 - яичники, 3 - желточники, 4 - семепроводы 5 - совокупительный орган, 6 - женское половое стверстие, 7 - яйцеводы
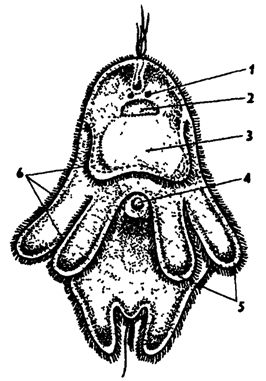
Рис. 121. Мюллеровская личинка (из Байера): 1 - глаза, 2 - ганглий, 3 - кишечник, 4 - рот, 5 - лопасти, 6 - мерцательный венчик
168
(рис. 120). У наиболее распространенных трехветвистых планарий половая система имеет следующее строение.
Мужская система представлена двумя семенниками, состоящими из семенных мешочков с семявыносящими канальцами. Парные семяпроводы впадают в семяизвергательный канал, пронизывающий совокупительный орган - пенис, который открывается в половую клоаку - кожное впячивание, куда открываются и женские протоки. В некоторых случаях семяизвергательный канал образует расширение - семенной пузырек, в котором накапливается сперма. Нередко имеются предстательные железы, впадающие в семенной пузырек.
Женская половая система представлена парными яичниками и отходящими от них яйцеводами. На яйцеводах расположены многочисленные железки - желточники, продуцирующие желточные клетки, переполненные желтком, необходимым для питания развивающихся яиц. Желточники представляют собой видоизмененные части яичника, а желточные клетки гомологичны ооцитам. Яйцеводы впадают во влагалище, открывающееся половым отверстием в клоаку. У некоторых видов в половую клоаку впадает еще и копулятивная сумка с семяприемником, куда после спаривания попадает сперма партнера. Оплодотворение яиц происходит позднее, после спаривания.
Оплодотворенная яйцеклетка вместе с группой желточных клеток покрывается скорлупой, и образуется сложное яйцо, характерное для большинства плоских червей.
Наиболее примитивное состояние половой системы у бескишечных турбеллярий. У них отсутствуют половые протоки, желточники. При спаривании бескишечных планарий сперма вводится в тело партнера при помощи копулятивного органа в паренхиму, где и происходит оплодотворение яиц, созревающих в паренхиме. Яйца простые (без желточных клеток), выпадают из тела в воду через разрывы стенки тела.
Размножение, развитие. Развитие у некоторых морских турбеллярий (Polycladida) происходит с метаморфозом, и из яйца выходит мюллеровская личинка (рис. 121). Она имеет овальную форму, покрыта ресничками и снабжена восемью радиально расположенными лопастями, помогающими парить в толще воды. На вершине тела личинки имеется рот, ведущий в мешковидный кишечник.
У большинства видов развитие прямое. Встречаются виды планарий, способные размножаться бесполым путем. При этом особь делится поперечными перетяжками на несколько дочерних особей.
В настоящее время выделяются два подкласса турбеллярий: подкласс Архоофоры (Archoophora) и подкласс Неоофоры (Neoophora), включающие 12 отрядов. Рассмотрим некоторые из них.
169
Подкласс Архоофоры (Archoophora)
Это наиболее примитивные планарии, у которых нет желточников, яйца простые, развитие нередко с метаморфозом. Имеется статоцист.
Отряд Бескишечные (Acoela). У них отсутствуют кишечник, протонефридии, половые протоки, желточники. Имеется мезодермальная и энтодермальная паренхима. Пища заглатывается ртом и переваривается в энтодермальной паренхиме. Половые клетки созревают в мезодермальной паренхиме. Нервная система - примитивный ортогон с диффузной сетью нервных тяжей. Это мелкие морские обитатели дна литоральной зоны.
Отряд Многоветвистые (Polycladida). Кишечник многоветвистый, множественность половых желез, отсутствие желточников. Развитие с метаморфозом. Это крупные морские формы (до 15 см).
Подкласс Неоофоры (Neoophora)
Эволюционно продвинутая группа планарии, перешедших к обитанию в пресных водах и даже на суше. У них имеются желточники. Яйца сложные. Развитие прямое.
Отряд Трехветвистые (Tricladida) - самый многочисленный по числу видов. Преимущественно пресноводные формы, реже морские. Некоторые тропические виды планарии - наземные обитатели. В наших пресных водоемах часто встречаются такие крупные планарии, как молочно-белая (Dendrocoelum lacteum) длиной 2-3 см и многоглазка (Polycelis, рис. 117). У них трехветвистый кишечник, хорошо развиты желточники, сложная половая система. Яйца сложные. Развитие прямое.
Отряд Прямокишечные (Rhabdocoela) - мелкие формы, встречающиеся в морях, пресных водах и во влажном мху на суше. Среди них имеются и паразитические виды. У них рот на переднем конце тела, прямой кишечник, хорошо развиты продольные стволы нервной системы, имеется сложная половая система с желточниками. Развитие прямое.
Происхождение турбеллярий. Турбеллярии - наиболее примитивная группа плоских червей, поэтому проблема их филогенетического происхождения связана и с эволюцией всего типа Plathelminthes. Имеется несколько гипотез происхождения плоских червей, которые отличаются тем, что в качестве плезиоморфных групп среди ныне живущих турбеллярий рассматривались разные отряды
По гипотезе Ланга, к самым примитивным турбелляриям относятся многоветвистые (Polycladida), которые якобы сохранили элементы радиальной
170
симметрии от предковой группы Radiata. Среди Radiata вьщеляли ползающих гребневиков, которые сочетали черты радиальной симметрии с признаками билатеральных животных. Обращалось внимание на такие черты сходства между гребневиками и многоветвистыми планариями: ресничный покров, строение пищеварительной системы с радиальными ветвями средней кишки, наличие статоциста над мозговым ганглием. Однако сравнительное изучение эмбрионального развития турбеллярий и гребневиков доказало несостоятельность этой гипотезы. Выяснилось, что поликладиды - весьма продвинутая группа турбеллярий, а наиболее примитивны бескишечные турбеллярий.
Вторая гипотеза происхождения турбеллярий была предложена Граффом, а позднее поддерживалась В. Н. Беклемишевым. К плезиоморфным турбелляриям были отнесены бескишечные (Acoela), которых выводили от планулообразных предков Radiata. Но Acoela не имеют оформленного кишечника, и поэтому их нельзя было считать потомками взрослых фаз кишечнополостных и гребневиков со сложной кишечной полостью. Авторы гипотезы допускали, что личинки предковых Radiata перешли через неотению (укорочение онтогенеза) к образованию бескишечных турбеллярий. Однако личинки современных кишечнополостных не питаются и представляют упрощенные формы в связи с образованием расселительной фазы. Если допустить происхождение плоских червей от планулоподобных, то нужно признать регресс от Radiata к Bilateria в связи с редукцией кишечника и радиально-симметричных органов.
Наиболее аргументирована гипотеза происхождения турбеллярий от фагоцителлоподобных предков. А. В. Иванов предложил гипотезу независимого происхождения Radiata и Bilateria от фагоцителлоподобных предков. Описание трихоплакса как единственного современного представителя Phagocytellozoa показывает, что такие формы очень близки к предкам бескишечных турбеллярий: ресничный покров, паренхиматозность. Ароморфоз в происхождении турбеллярий заключался в том, что паренхима фагоцителлоподобных предков расслоилась на два функциональных пласта: внутренний - фагоцитобласт, в котором осуществлялось внутриклеточное пищеварение, и промежуточный - мезодермальную паренхиму, имеющую опорное значение. У высших планарий в мезодерме формируется и мускулатура. Погружение фагоцитобласта внутрь тела привело к образованию рта, расположенного на нижней поверхности тела. А это, в свою очередь, повлекло образование билатеральной симметрии. Эта гипотеза без натяжек объясняет простоту организации Acoela и прогресс по сравнению с фагоцителлообразными. Из гипотезы вытекает вывод о разных путях специализации Radiata и Bilateria. Согласно этой системе взглядов, гребневики и поликладиды -
171
одни из самых специализированных групп в разных ветвях эволюции (Radiata и Bilateria) и потому не могут быть связаны непосредственным родством. От бескишечных турбеллярий эволюция шла по пути усложнения организации: пищеварительной, нервной, выделительной и половой систем органов.
Особняком стоит гипотеза Хаджи о происхождении турбеллярий от инфузорий, о которой уже упомянуто в связи с изложением состояния проблемы о происхождении многоклеточных. По Хаджи, в процессе эволюции от полиэнергидных инфузорий путем целлюляризации могли возникнуть первичные бескишечные турбеллярий с наружным ресничным эпителием и внутренней паренхимой. Если принять эту концепцию, то следует признать, что первичными многоклеточными были трехслойные животные - планарии, от которых в дальнейшем произошли такие более примитивные типы, как губки и кишечнополостные, что не соответствует постулатам учения о зародышевых листках и принципах онтофилогенеза.
Поэтому наиболее аргументированной гипотезой происхождения турбеллярий в настоящее время следует считать концепцию Иванова о фагоцителлоподобном предке первых билатеральных животных и ацелоподобном предке плоских червей. Согласно этой концепции можно следующим образом представить происхождение низших многоклеточных и первичных билатеральных животных (рис. 122).
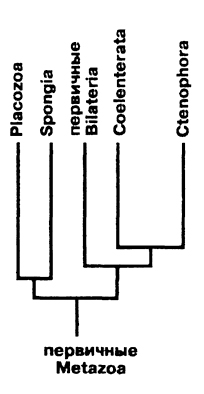
Рис. 122. Филогенетические отношения Bilateria с другими Metazoa
172
3W.SU ©® 2015