Зоология беcпозвоночных
Филогения и экологическая радиация кольчатых червей
В разных классах кольчатых червей прослеживается ряд плезиоморфных признаков, свидетельствующих об их родстве с низшими червями (Acoelomata): наличие первичной полости тела, протонефридиев у примитивных представителей и у личинок, аметамерность или олигомерность строения, ресничный тип движения, отсутствие кровеносной системы.
Так, у примитивных Archiannelida и Polychaeta вторичная полость может быть неразвита, нередко отсутствует кровеносная система, но имеются первичная полость тела, протонефридии, характерные для первичнополостных червей. Нервная система, близкая к ортогональной, наблюдается
270
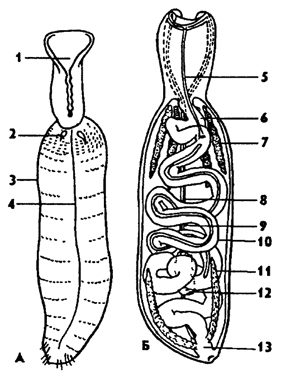
Рис. 191. Эхиуриды Echiurus (по Барнсу): А - внешнее строение с вентральной стороны, Б - внутреннее строение (с дорсальной стороны); 1 - желобок, 2 - щетинка, 3 - туловище, 4 - брюшной нервный ствол, 5 - кишечный сосуд, 6 - передняя кишка, 7 - нефридий, 8 - нервный ствол, 9 - вентральный кровеносный сосуд, 10 - средняя кишка, 11 - анальный мешок, 12 - гонада, 13 - прямая кишка
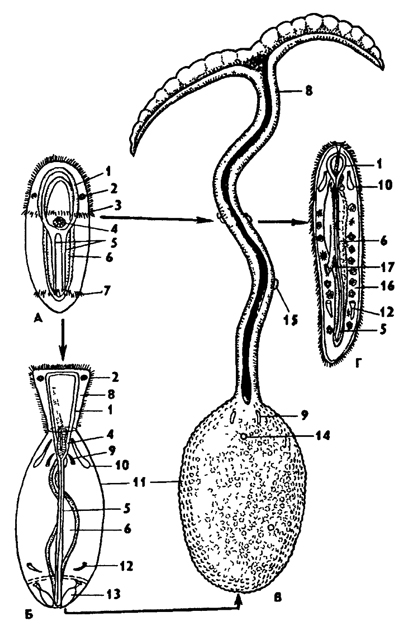
Рис. 192. Бонелия Bonellia vindis (по Бальтцеру): А - личинка, Б - женская личинка старшего возраста, В - взрослая самка, Г - самец; 1 - окологлоточное нервное кольцо, 2 - глазное пятно, 3 - прототрох, 4 - передняя кишка, 5 - брюшные нервные стволы, 6 - полость средней кишки, 7 - анальный венчик ресниц, 8 - хоботок, 9 - щетинки, 10 - протонефридий, 11 - тело, 12 - метанефридий, 13 - анальный мешок с воронкой, 14 - женское половое отверстие, 15 - личинка на хоботке самки, 16 - целом с семенными клетками, 17 - семенной проток
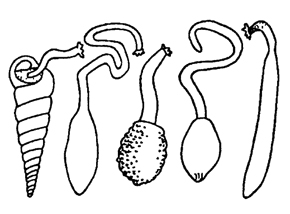
Рис. 193. Форма тела у сипункулид (по Барнсу)
271
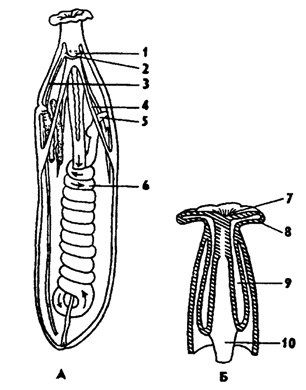
Рис. 194. Внутреннее строение сипункулиды (по Барнсу): А - вскрытая сипункулида, Б - передний конец тела с гидравлической системой щупалец; 1 - мозг, 2 - нервное кольцо, 3 - вентральный нервный ствол, 4 - ретрактор 5 - задняя кишка, 6 - средняя кишка, 7 - рот, 8 - щупальца, 9 - щупальцевый мешок, 10 – пищевод
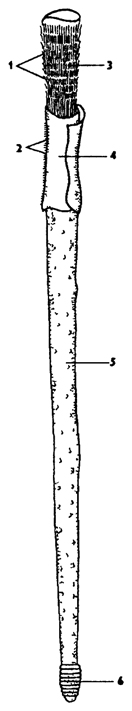
Рис. 195. Погонофоры - вестиментифера (по Барнсу): 1 - жабры, 2 - реснички на жабрах, 3 - обтуракулум, 4 - железистый отдел, 5 - туловище, 6 - задний кольчатый отдел
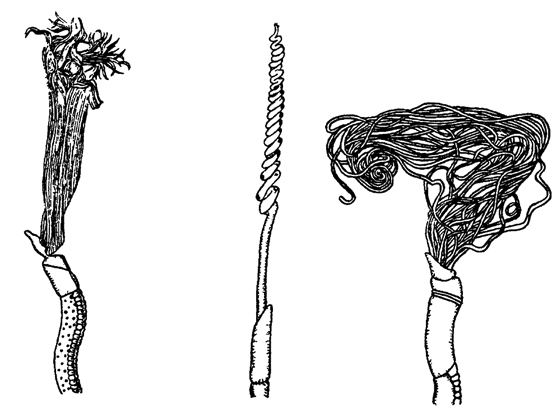
Рис. 196. Передние концы тела с жабрами трех видов погонофор - первиат (по Иванову)
272
у личинки аннелид - трохофоры, а также у сипункулид и эхиурид. У трохофоры на ранней фазе развития может быть слепозамкнутый кишечник, как у плоских червей. Самцы Bonellia из эхиурид похожи на планарий: покрыты ресничным эпителием и обладают слепозамкнутым кишечником. Аннелиды, как и круглые черви, сходны с прямокишечными турбелляриями по положению глотки на переднем конце тела.
Метамерия Annelida формируется в пределах типа. Так, эхиуриды и сипункулиды аметамерны, а у примитивных архианнелид и полихет метамерия затрагивает лишь эктодермальные производные (ларвальная сегментаци, по П. П. Иванову). Только у высших представителей типа (большинство Polychaeta, а также Oligochaeta и Hirudinea) метамерия касается не только эктодермальных, но и мезодермальных производных (постларвальная сегментация).
Целом, как оказалось, свойствен не всем кольчатым червям и возникает в разных группах по-разному: не только телобластически из мезодермальных зачатков, возникающих из клетоктелобластов, как описано выше, но и энтероцельно из первичной кишки, а также из схизоцеля, или из мускульного зачатка - миоцеля, или из половых желез (гоноцеля) (В. Н. Беклемишев). У одних групп кольчецов целом целостный (Sipunculida, Echiurida и некоторые Archiannelida), а у большинства - метамерный, с перегородками.
Предполагается, что гипотетические предки кольчецов были мелкими олигомерными или амерными (несегментированными) животными с цельным целомом и ресничным типом движения. Из современных аннелид наиболее близки к гипотетическим предкам некоторые архианнелиды, как Dinophilus, а также личинки полихет - метатрохофоры.
Филогенез Annelida в свете современных воззрений представляется следующим образом. От гипотетических предков аннелид прослеживается несколько путей эволюции. Один путь - это развитие полимерности строения в связи с укрупнением тела и развитием подвижности за счет мускулатуры. На раннем этапе этого процесса обособился класс Archiannelida, а на позднем - класс Polychaeta. Возникновение совершенной полимерной метамерии у полихет привело этот класс к широкой экологической радиации. От Polychaeta при переходе к пресноводному и наземному образу жизни сформировался класс малощетинковых червей - Oligochaeta, которые сохраняют множество общих черт организации с полихетами. При переходе к активному хищничанию и кровососанию от малощетинковых обособился класс пиявок Hirudinea, примитивные представители которых очень близки к олигохетам. Выход малощетинковых и пиявок в процессе эволюции в пресные воды и на сушу сопровождался переходом к прямому развитию. У олигохет развились адаптации к роющему
273
образу жизни, а у пиявок к эктопаразитизму и хищничеству. Филогенетические отношения классов Annelida отражены на рисунке 197.
Второй путь эволюционного развития Annelida связан с сохранением аметамерности строения и развитием приспособления к малоподвижному и неподвижному образу жинзи на морском дне, что привело к формированию классов Echiurida и Sipunculida. Однако эти классы развивались, по-видимому, независимо от разных аметамерных предков, а наблюдаемое у них сходство - лишь проявление конвергенции (мешковидное тело с хоботком).
Морфоэкологическая эволюция Annelida отражает экологические аспекты филогенеза. Центр экологической радиации кольчатых червей, несомненно, в морской среде, где обитает большинство представителей систематических групп кольчецов и наблюдается наибольшее многообразие жизненных форм. Наиболее примитивным адаптивным обликом обладают архианнелиды (типа tinophilus), представляющие собой донных эпибионтов со скользящим типом движения при помощи ресничек. От таких форм прослеживается путь специализации к полимерным донным эпибионтам с ползающим типом движения за счет сокращения мускулатуры и параподий (типа Phyllodoce). Донные ползающие формы, по-видимому, дали начало трем направлениям специализации. Одни полихеты перешли к роющему образу жизни с образованием роющих интрабионтов (типа Arenicola); другие перешли к плавающему образу жизни (пелагобионты); третьи приспособились к сидячему образу жизни (неподвижные бентобионты). У сидячих форм тело нередко заключено в трубку и на переднем конце тела сильно развиты жабры.
При переходе аннелид к роющему образу жизни в пресных водах сформировался класс малощетинковых червей, которые по облику похожи на морских роющих полихет - пескожилов. Малощетинковые претерпели менее разнообразную экологическую радиацию. Важнейшие направления в их специализации связаны, с одной стороны, с переходом к сидячему образу жизни (трубочники) и с выходом на сушу с образованием роющих геобионтов (дождевых червей). От пресноводных малощетинковых, вероятно, произошли плавающие хищные и эктопаразитические пиявки, некоторые из которых также вышли на сушу (почвенные и древесные пиявки в тропиках). Экологическая радиация кольчатых червей и близких к ним групп отражена на рисунке 198.
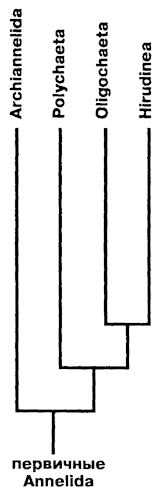
Рис. 197. Филогения кольчатых червей
274
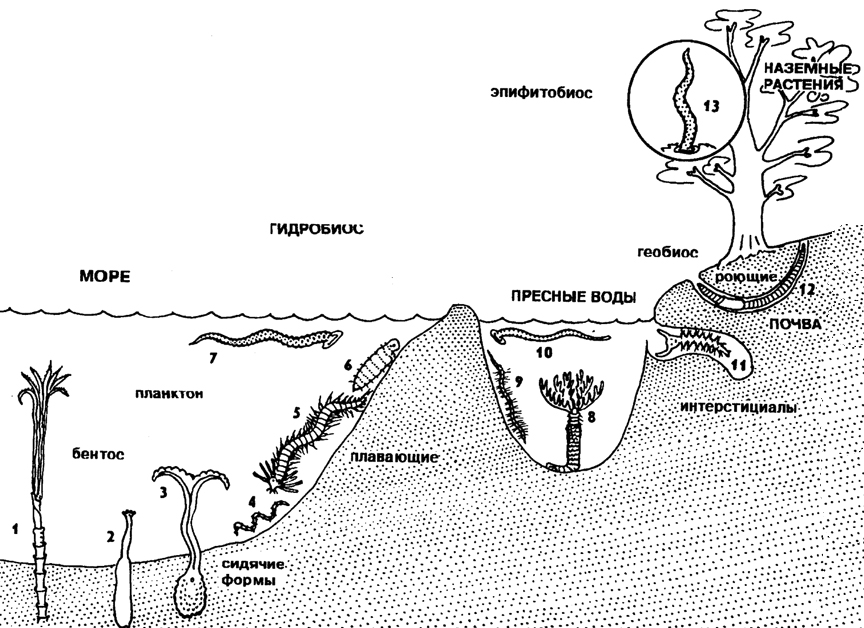
Рис. 198. Морфоэкологическая эволюция кольчатых червей и близких к ним типов: 1 - погонофора, 2 - сипункулида, 3 - бонелия, 4 - олигохета, 5 - полихета, 6 - архианнелида, 7 - пиявка морская, 8 - манаюнкия (полихета), 9 - олигохета, 10 - пиявка, 11 - троглохетус (пещерная полихета), 12 - дождевой червь, 13 - древесная пиявка
3W.SU ©® 2015